1
AMINOKWASY
zwi
ą
zki dwufunkcyjne;
w naturze > 500, kodowane – ok. 20
→
100 000 ró
ż
nych białek
Inne podziały:
•
Ze wzgl
ę
du na pozycj
ę
grupy aminowej (
αααα
-,
ββββ
-,
γγγγ
-….aminokwasy)
NH
2
-CHR-(CH
2
)
x
-COOH x = 0,1,2…; rz
ę
dowo
ść
gr. aminowej;
•
glikogenne (przetwarzane w glukoz
ę
lub glikogen w przypadku niedoboru
w
ę
glowodanów: Gly, Ala, Ser, Thr, Val, Asp, Glu, Arg, His, Met, Pro), ketogenne (keton
ko
ń
cowym produktem przemiany np. Leu)
•
Alifatyczne, aromatyczne, heterocykliczne…
•
Oboj
ę
tne, kwasowe, zasadowe
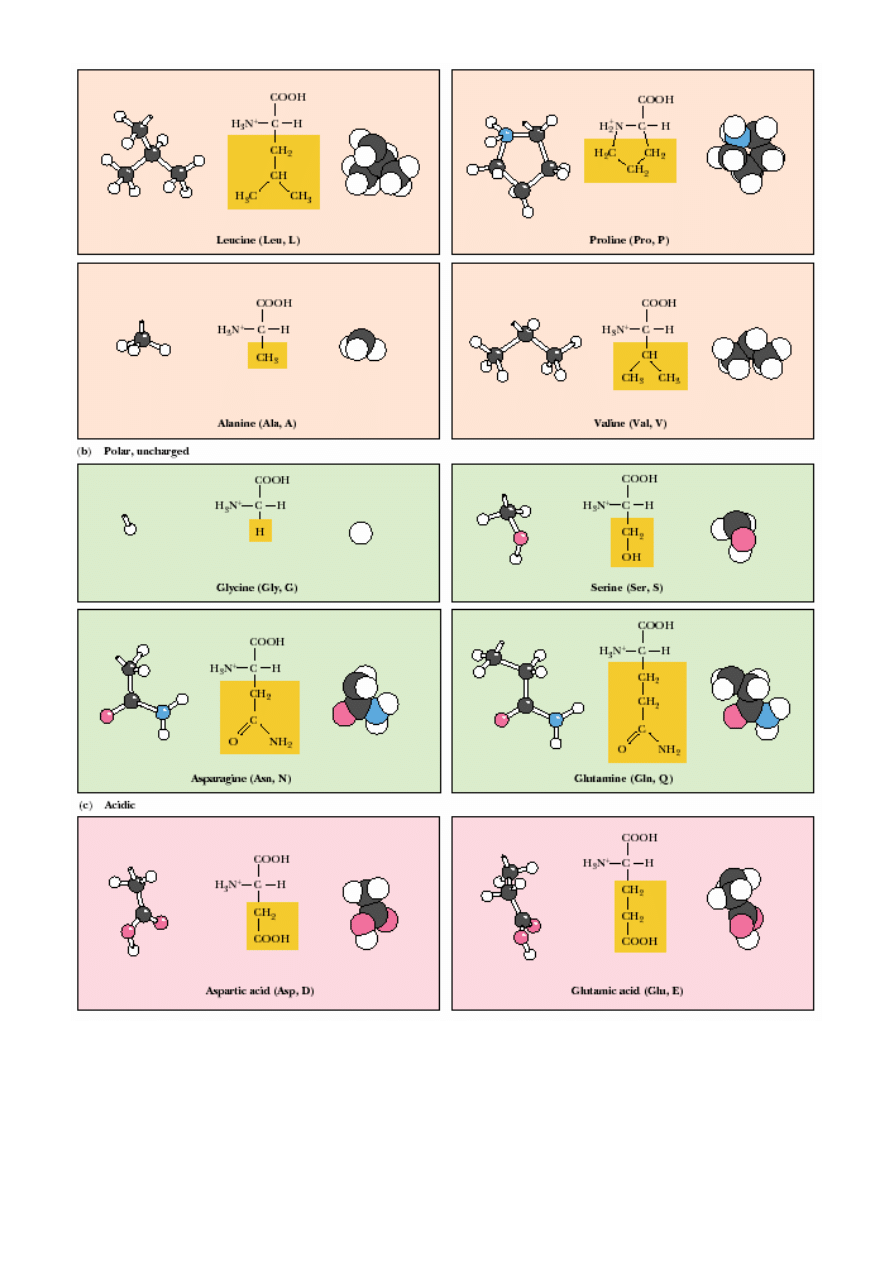
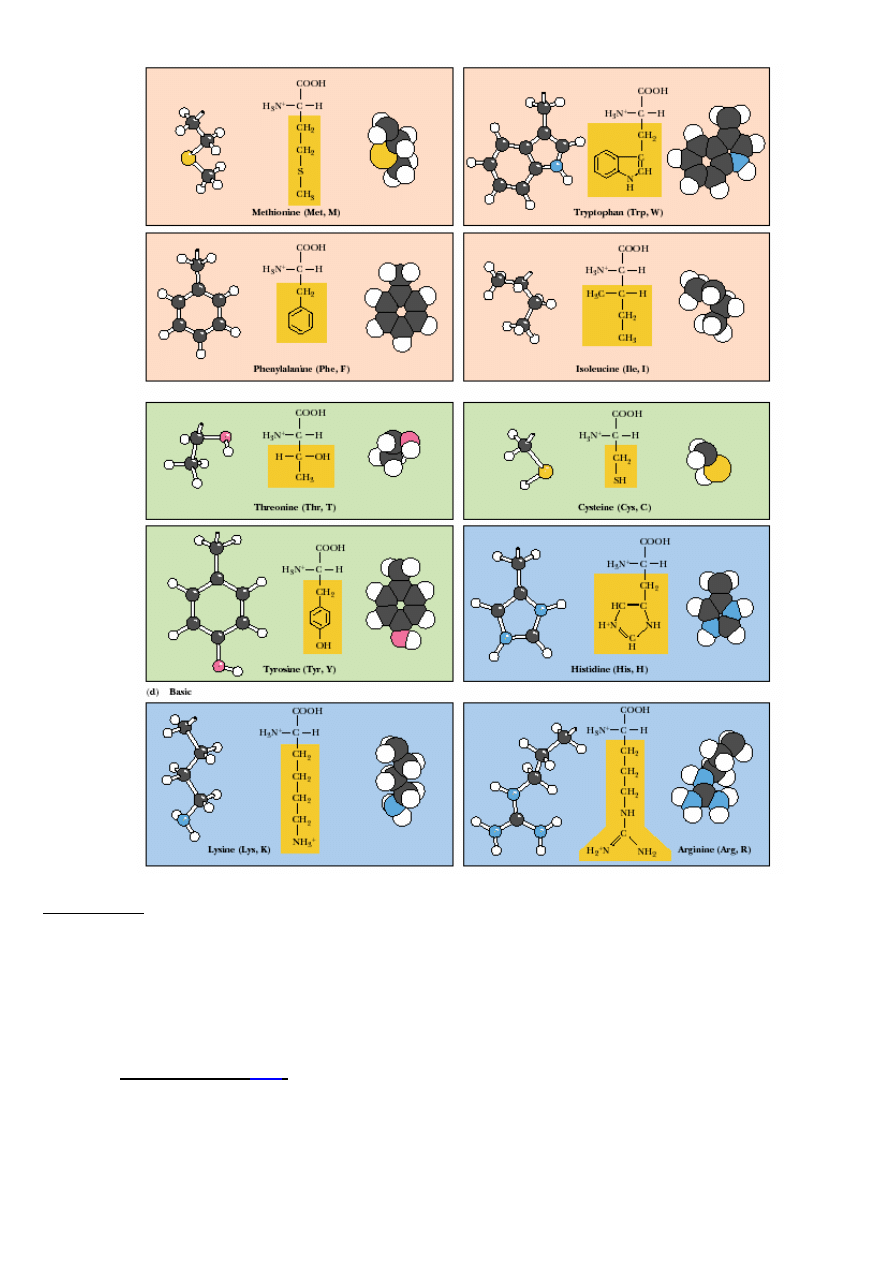
AMINOKWASY KODOWANE:
a/ niepolarne, hydrofobowe (łososiowy); b/ polarne nienaładowane (zielony); c/ kwasowe (ró
ż
);
d/ zasadowe (niebieski)
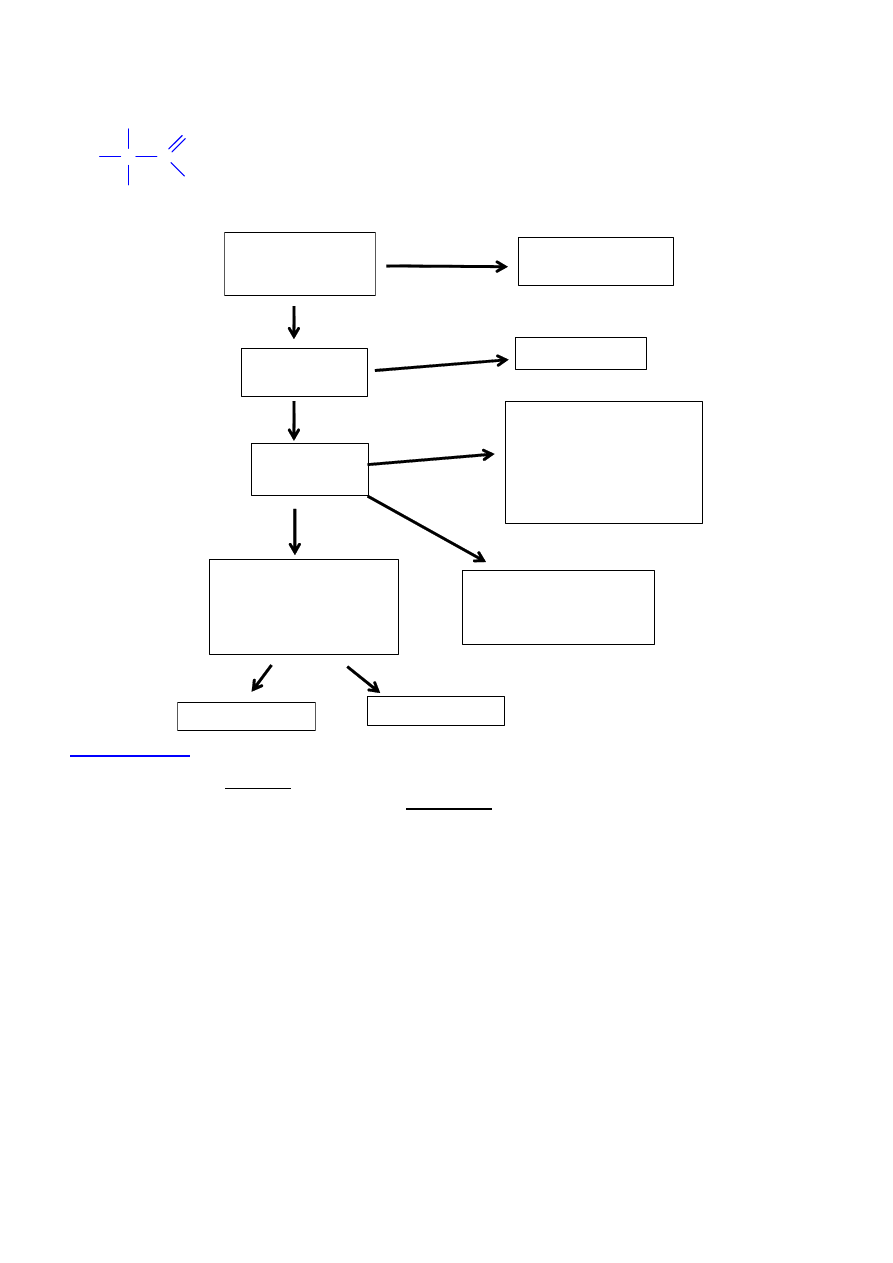
AMINOKWASY
NATURALNE
kilkaset
SYNTETYCZNE
(NIENATURALNE)
BIAŁKOWE
(kilkadziesi
ą
t)
NIEBIAŁKOWE
PIERWSZORZ
Ę
DOWE
(kodowane, 20 lub 21)
substraty w syntezie
rybosomalnej
DRUGORZ
Ę
DOWE
wynik postrybosomalnej
modyfikacji
TRZECIORZ
Ę
DOWE
modyfikacje
postrybosomalne z
niepeptydowymi
wi
ą
zaniami
kowalencyjnymi
ENDOGENNE
EGZOGENNE
C
O
OH
C
NH
2
R
H
aminokwas
α−
2
3
Aminokwasy:
- niepolarne, hydrofobowe, np.:
Leu
,
Ile,
Pro,
Val
, Ala,
Trp
,
Phe
,
Met
…
- polarne, nienaładowane, np.: Gly, Ser, Asn, Cys,
Thr
, Tyr…
- kwa
ś
ne, np.: Asp, Glu;
- zasadowe, np.:
Arg
,
Lys
,
His
.
20 Aaa kodowanych,
8 Aaa
niezb
ę
dnych
(nie syntezowanych przez dojrzałego człowieka)
21 Aaa – selenocysteina (
Sec
) NH
2
-CH-COOH
1986 r.
|
CH
2
-Se
rekodowanie kodonu terminalnego UGA; Sec bardziej nukleofilowa vs Cys
→
enzymy

4
22 Aaa – L-pyrrolysine
B. Hao, W. Gong, T.K.Ferguson, C.M. James, J.A. Krzycki, M.K. Chan, A new UAG-encoded residue
in the structure of a mathanogen methyltransferase, Science 2002, 296, 1462
N
N
H
OH
O
O
NH
2
X
X = OH, NH
2
,CH
3
“Stop codon” UAG enzymatycznie reprogramowany do syntezy “pyrrolysine”. Nieoczekiwana
ró
ż
norodno
ść
⇒
nowe enzymy o znaczeniu przemysłowym.?
Hydroliza białek
→
24 Aaa (dawniej - 22 Aaa) (kodowane + Cys-Cys + Hyp)
2R
SH
[O]
[H]
R
-S-S-
R
Nomenklatura i symbolika:
-
nazwa zwi
ą
zana z: pierwsz
ą
izolacj
ą
[
ź
ródłem - Asp (łac. – asparagus), Cys (gr. – cystis –
p
ę
cherz], sposobem wyodr
ę
bniania (Arg – w post. soli Ag, Trp – prod. degr. trypsyn
ą
),
podobie
ń
stwem do innych zwi
ą
zków (Val – od kw. walerianowego)
-
z wyj
ą
tkiem Trp, przyrostek
–ina lub –yna
-
reszty
(H
2
N-CHR-CO-) przyrostek
–ylo
[glutamylo- (Glu-), aspartylo-, glutaminylo- (Gln-),
asparaginylo-]
-
skróty trzyliterowe, konfiguracja w
ę
gla
α
zawarta w symbolu (L-alanina = Ala, A –
jednoliterowy skrót biochemiczny)
COOH
H
R
H
2
N
COOH
H
CH
2
OH
HO
L-aminokwas
aldehyd L-glicerynowy
(S)-2,3-dihydroksypropanal
-
allo
-L-izoleucyna – aIle,
hydroksy
lizyna –
Hy
l,
allo
-L-
hydroksy
prolina -
aHy
p
-
nor
walina –
N
va
-
grupa aminowa – A (kw.
α
-aminomasłowy – Abu,
α
-aminoadypinowy Aad)
dwie grupy aminowe – D (
α
,
γ
-diaminomasłowy – Dbu)
poło
ż
enia w pozycjach
α
i
ω
nie s
ą
uwzgl
ę
dniane w nazwie, inne tak (
β
-Ala)
N-metylowalina – MeVal
WŁA
Ś
CIWO
Ś
CI CHEMICZNE
Aminokwasy s
ą
zwi
ą
zkami amfoterycznymi, tworz
ą
sole obojnacze.
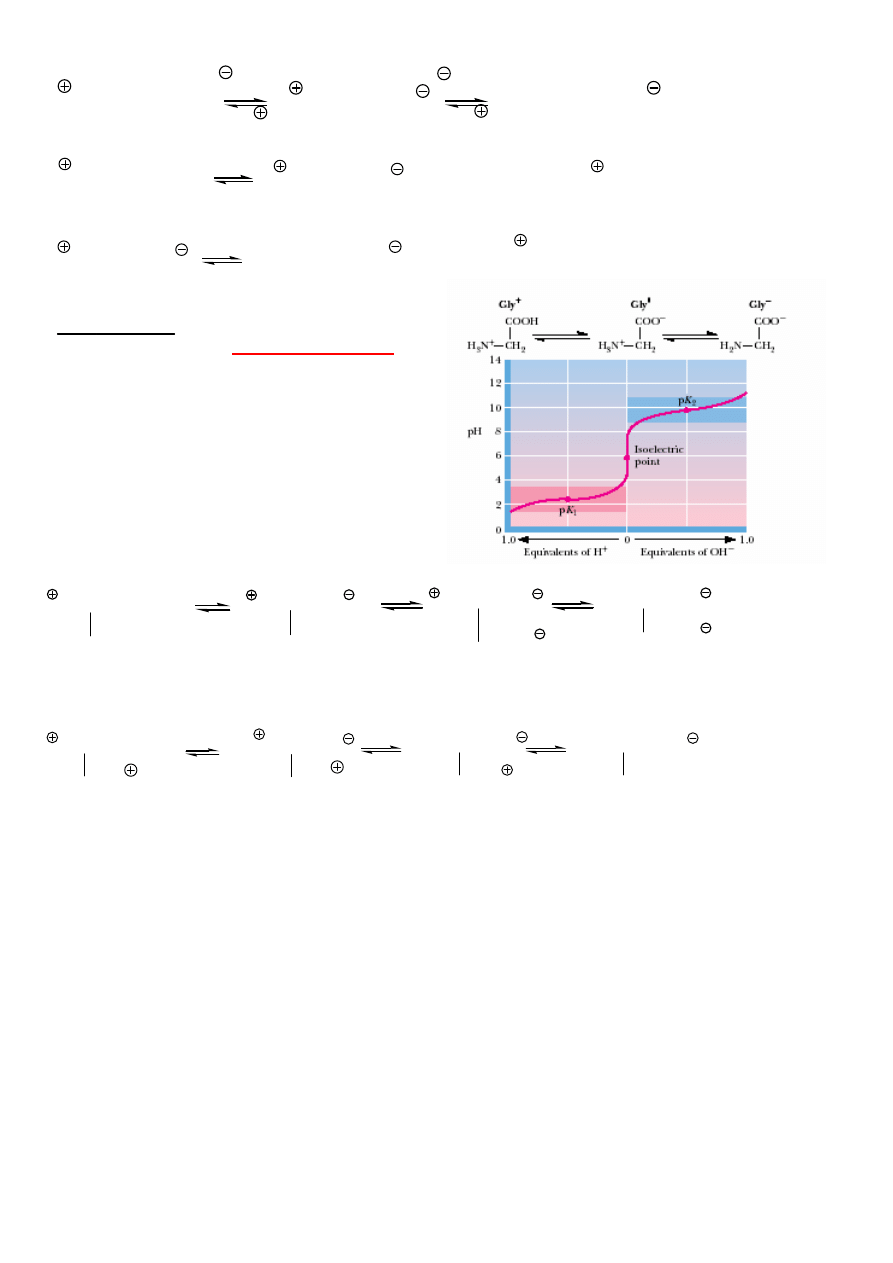
5
NH
3
-CHR-COOH
NH
3
-CHR-COO
NH
2
-CHR-COO
OH
OH
H
H
NH
3
-CH
2
-COOH
NH
3
-CH
2
-COO
pK
a,1
= 2.4 (wplyw NH
3
)
K
a,1
NH
3
-CH
2
-COO
NH
2
-CH
2
-COO
pK
a,2
= 9.8 (NH
3
, a nie COOH)
K
a,2
punkt izoelektryczny
max. st
ęż
enie jonu obojnaczego (minimalny
ładunek)
Gly - pI = ½ (2.4 + 9.8) = 6.1
CH
2
COOH
NH
3
-CH-COOH
CH
2
COOH
NH
3
-CH-COO
CH
2
COO
NH
3
-CH-COO
pK
1
1.9
pK
2
CH
2
COO
NH
2
-CH-COO
pK
3
3.7
9.6
pI
= ½ (1.9 + 3.7) = 2.8 (Asp)
(CH
2
)
4
NH
3
NH
3
-CH-COOH
(CH
2
)
4
NH
3
H
3
N-CH-COO
(CH
2
)
4
NH
3
NH
2
-CH-COO
pK
1
2.2
pK
2
(CH
2
)
4
NH
2
NH
2
-CH-COO
pK
3
9.0
10.5
pI
= ½ (9.0 + 10.5) = 9.7 (Lys)
Ż
RÓDŁA AMINOKWASÓW
1. Hydrolizaty białkowe
2. Metody mikrobiologiczne
3. Metody enzymatyczne
•
rozdział racemicznych pochodnych Aaa
•
synteza asymetryczna
4. Metody syntetyczne
→
produkty achiralne/produkty chiralne
pK
a,1
+ pK
a,2
2
= pI
6
SYNTEZA
1. Bromowanie i aminowanie kwasów karboksylowych
CH
3
CH
2
COOH
Br
2
PBr
3
CH
3
CHCOOH
Br
CH
3
CHCOOH
NH
2
NH
3,
H
2
O
(R,S)-Ala, 56%
80%
Małe zastosowanie, zwykle niskie wydajno
ś
ci.
2. Synteza Gabriela
(z ftalimidku potasu)
Alkilowanie anionu ftalimidowego (1,2-benzenodikarboksylowego). Czynnik alkiluj
ą
cy – produkt
bromowania malonianu dietylowego.
N K
O
O
+
HC
COOC
2
H
5
COOC
2
H
5
Br
N
O
O
CH
COOC
2
H
5
COOC
2
H
5
N
O
O
CR
COOC
2
H
5
COOC
2
H
5
1. EtO Na, EtOH
2. RX
H, T
N
O
O
CR
COOH
COOH
-CO
2
N
O
O
CHR
COOH
COOH
COOH
NH
3
-CHR-COO
ester imidomalonowy
3. Synteza Strecker’a
Działanie amoniakiem i cyjanowodorem na aldehydy.
R
C
O
H
C
OH
NH
2
H
R
C
NH
R
H
R
C
NH
2
CN
H
NH
3
-H
2
O
HCN
H, H
2
O, T
R-HC-COOH
NH
2
aminonitryl
4. Redukcyjne aminowanie
αααα
-oksokwasów
(laboratoryjny analog biosyntezy)
H
3
C
C
O
COOH
NH
3
NaBH
4
H
3
C
H
C
COOH
NH
2
R,S-Ala
kwas
αααα−−−−
oksopropanowy
(pirogronowy)
7
5. Synteza amidomalonianowa
Najbardziej uniwersalna.
C
COOEt
COOEt
NH-CO-CH
3
C
COOEt
COOEt
NH-CO-CH
3
R
R-X
S
N
2
EtONa
C
COOEt
COOEt
NH-CO-CH
3
H
H
3
O
H
3
O
+ CO
2
+ 2EtOH + CH
3
COOH
C
COOEt
COOEt
NH-CO-CH
3
C
COOEt
COOEt
NH-CO-CH
3
R
R-X
S
N
2
EtONa
C
COOEt
COOEt
NH-CO-CH
3
H
H
3
O
H
3
O
R-CH-COOH
NH
2
+ CO
2
+ 2EtOH + CH
3
COOH
6. Syntezy enancjoselektywne (asymetryczne)
Np.:
A.
U
ż
ycie chiralnych katalizatorów otrzymanych na bazie metali przej
ś
ciowych:
H
2
C
C
COOH
NHCOCH
3
C
COOH
H
3
COCHN
H
3
C
H
chiralny kompleks Rh
H
2
kwas 2-acetyloaminopropenowy
B.
Biosynteza – redukcyjne aminowanie
α
-oksokwasów:
HOOCCH
2
CH
2
CCOOH
O
kwas
αααα−−−−
ketoglutarowy
(
α
αα
α−−−−
oksoglutarowy)
+ NH
4
+ NADPH
dehydrogenaza
glutaminianowa
HOOCCH
2
CH
2
CHCOOH
NH
2
+ NADP + H
2
O
L-Glu
Escherichia coli – synteza wszystkich kodowanych; człowiek – prócz 9 egzogennych
L-Glu (prekursor)
→
Gln, Pro, Arg
Wi
ę
kszo
ść
Aaa – grupa
α
-aminowa
←
Glu
H
C
R
NH
3
COO
+ R'CCOOH
O
transaminaza
R'CHCOO
NH
3
RCOCOOH +
7. Rozdział racemicznych aminokwasów:
A.
za pomoca deacylaz:
8
NH
3
NHCOCH
3
Ac
2
O
deacylaza
H
3
N
C
R
H
COO
H
C
R
NHAc
COOH
+ CH
3
COOH
+
R-CH-COOH
R-CH-COO
B.
z u
ż
yciem chiralnych amin - (-)-chinina, (-)-strychnina, (-)brucyna lub kwasu (+) lub (-)
winowego
H
C
R
NH
3
COO
H
3
N
C
R
H
COO
Ph
C
R
H
NH
2
racemiczny Aaa
(R)-1-fenyloetyloamina
+
sól S,R
sól R,R
R-Aaa
S-Aaa
C.
HPLC z chiralnymi fazami stacjonarnymi (HPLC-CSP)
PEPTYDY
CH
C
N
H
CH
C
H
N
CH
C
OH
O
O
O
R"
R'
R
H
2
N
C-koniec
N-koniec
Aaa
1
Aaa
3
Aaa
2
Ła
ń
cuch główny, ła
ń
cuchy boczne (R, R’, R”)
Dipeptyd, tripeptyd…
Sekwencja Aaa w ła
ń
cuchu peptydowym:
H
3
N-H
2
C-C-NH-CH-COO
O
CH
3
H
3
N-HC-C-NH-CH
2
-COO
O
CH
3
Gly-Ala
Ala-Gly
SYNTEZA W ROZTWORZE
Gly + Ala
- H
2
O
∆
Gly-Gly + Ala-Gly +
Gly-Ala
+ Gly-Gly-Ala....
9
P
1
Gly + AlaOP
2
aktywacja
P
1
Gly-AlaOP
2
HGly-AlaOH
I. Osłona grupy aminowej (gł. grupy uretanowe):
A. Grupa benzyloksykarbonylowa (Z, Cbz):
C
6
H
5
CH
2
-O-C-Cl
O
+ NH
3
CHCOO
CH
3
NaOH
-NaCl
-HOH
C
6
H
5
CH
2
-O-C
O
CH
3
NHCHCOOH
chloromrówczan benzylu
(Z-Cl)
Z-Ala
C
6
H
5
CH
2
-O-C
O
NHCH
2
RCO--
H
2,
Pd
C
6
H
5
CH
3
+ CO
2
+ NH
2
-CHRCO-
C
6
H
5
CH
2
Br + CO
2
+ NH
3
-CHR-CO
-
HBr
B. Grupa tert-butoksykarbonylowa (Boc, BOC)
NH
3
-CHR-COO
+ (CH
3
)
3
CO-C-O- C-O-C(CH
3
)
3
O
O
diw
ę
glan tert-butylowy
B
-CO
2
-(CH
3
)
3
COH
(CH
3
)
3
COC
O
NHCHRCOOH
(Boc
2
O)
Zdejmowanie:
(CH
3
)
3
COC
O
NHCHRCO
HCl lub CF
3
COOH
H
3
NCHC
R O
+ CO
2
+ CH
2
=C(CH
3
)
2
C. Grupa fluorenylo-9-metoksykarbonylowa (Fmoc)
CH
2
O
CNH
O
C
H
R
COOH
NH
CH
2
x C
5
H
11
N
+ NH
2
-CH-COOH
R
Fmoc-Aaa
II. Osłona grupy karboksylowej: estry benzylowe, tert-butylowe, metylowe…
OBzl – H
2
/Pd
OtBu – TFA
OMe – NaOH
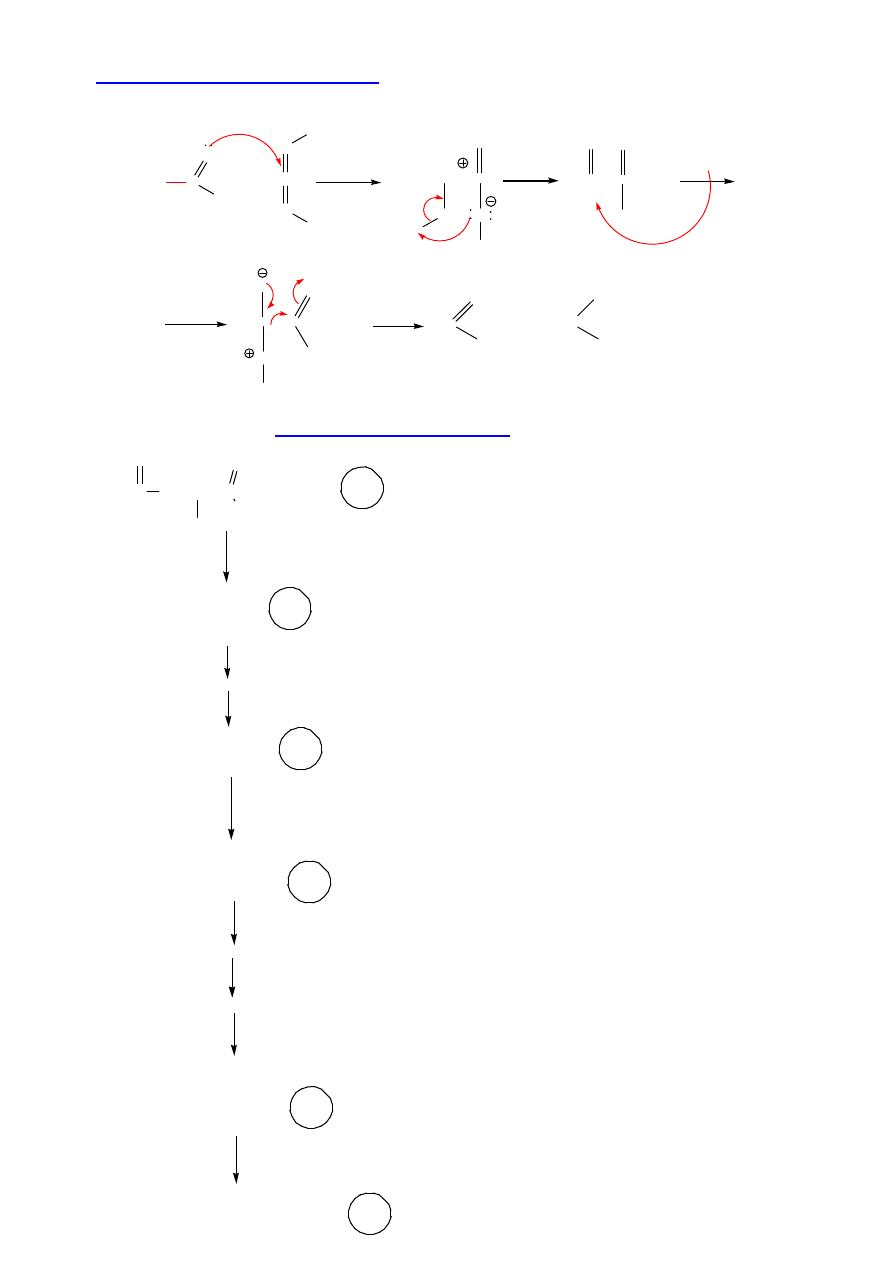
10
III. Aktywacja grupy karboksylowej:
Dicykloheksylokarbodiimid DCC, C
6
H
11
N=C=N-C
6
H
11
R
C
O
OH
N
C
N
C
6
H
11
C
6
H
11
+
R-C=O-C
N-C
6
H
11
N
C
6
H
11
O
H
R-C-O-C
NC
6
H
11
NHC
6
H
11
R"-NH
2
O
R-C-O-C
O
NH
2
NC
6
H
11
NHC
6
H
11
R'
R-C
O
NH-R'
+ O=C
NHC
6
H
11
NHC
6
H
11
DCU
Synteza na no
ś
niku stałym (Merrifield’a)
R. Bruce Merrifield – nN 1984 r.
Automatyczne syntezatory peptydów – 1 Aaa – ok. 1 h
Synteza rybonukleazy (124 Aaa) – 369 reakcji,
12 000 zautomatyzowanych kroków;
η
= 17%
(1 etap> 99%)
Biosynteza – 150 Aaa w okre
ś
lonej sekwencji 1 min (!)
(CH
3
)
3
C-O-C
O
NH-CH-C
O
OH
R
+ Cl-CH
2
- R
B
Boc-Aaa
1
R
-CH
2
-
TFA
R
3
N
NH
2
-Aaa
1
R
-CH
2
-
Boc-Aaa
2
OH
R
Boc-Aaa
2
-Aaa
1
-CH
2
-
TFA
Et
3
N
Boc-Aaa
3
OH
R
Boc-Aaa
3
-Aaa
2
-Aaa
1
-CH
2
-
HF
Aaa
3
-Aaa
2
-Aaa
1
-OH
+ F-CH
2
-
R
DCC
DCC
11
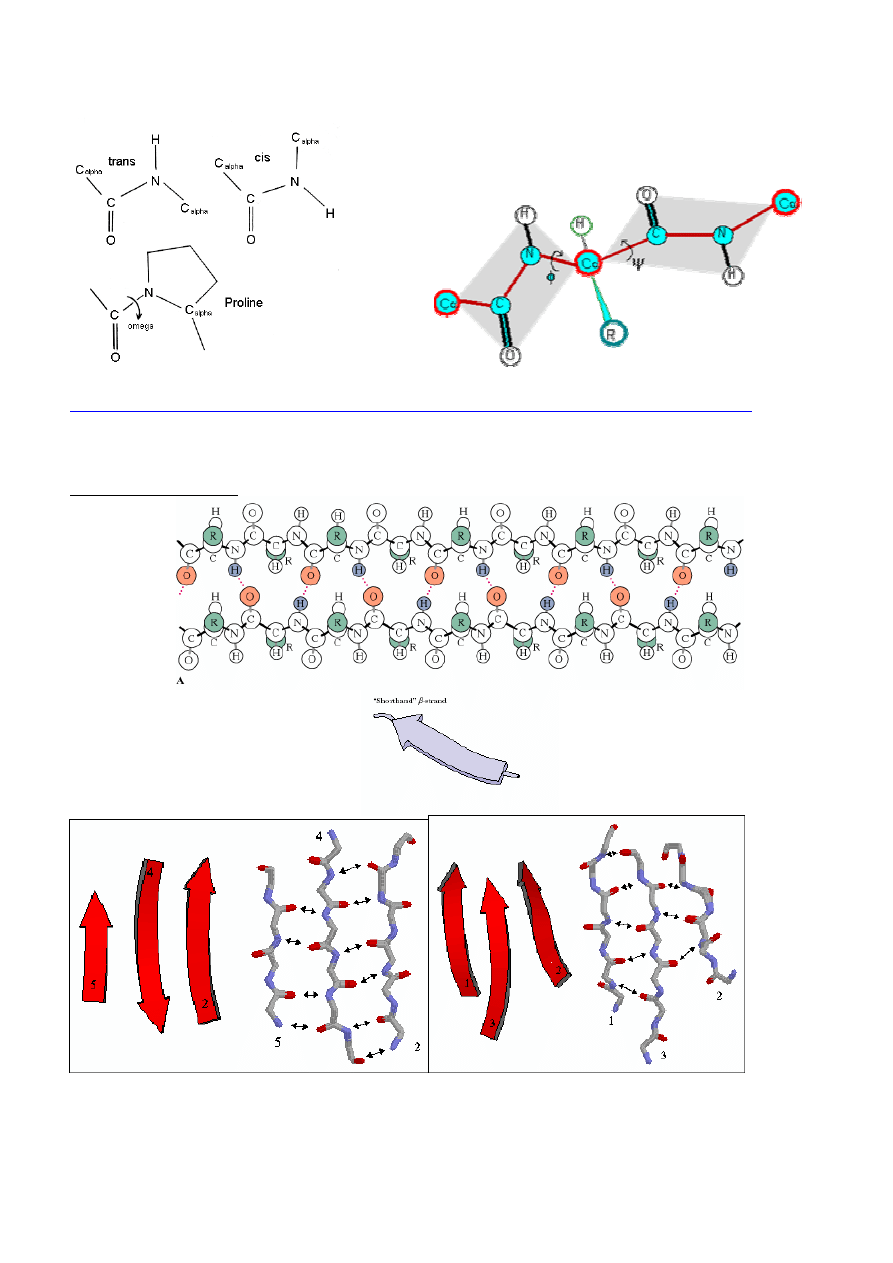
STRUKTURA POLIPEPTYDÓW /BIAŁEK
Sekwencja Aaa w ła
ń
cuchu peptydowym –
1° struktura peptydu/białka
Płaski układ wi
ą
zania peptydowego/amidowego:
C
C
O
N
H
HN
C
NH
O
R
H
H
R
C
C
O
N
H
HN
C
NH
O
R
H
H R
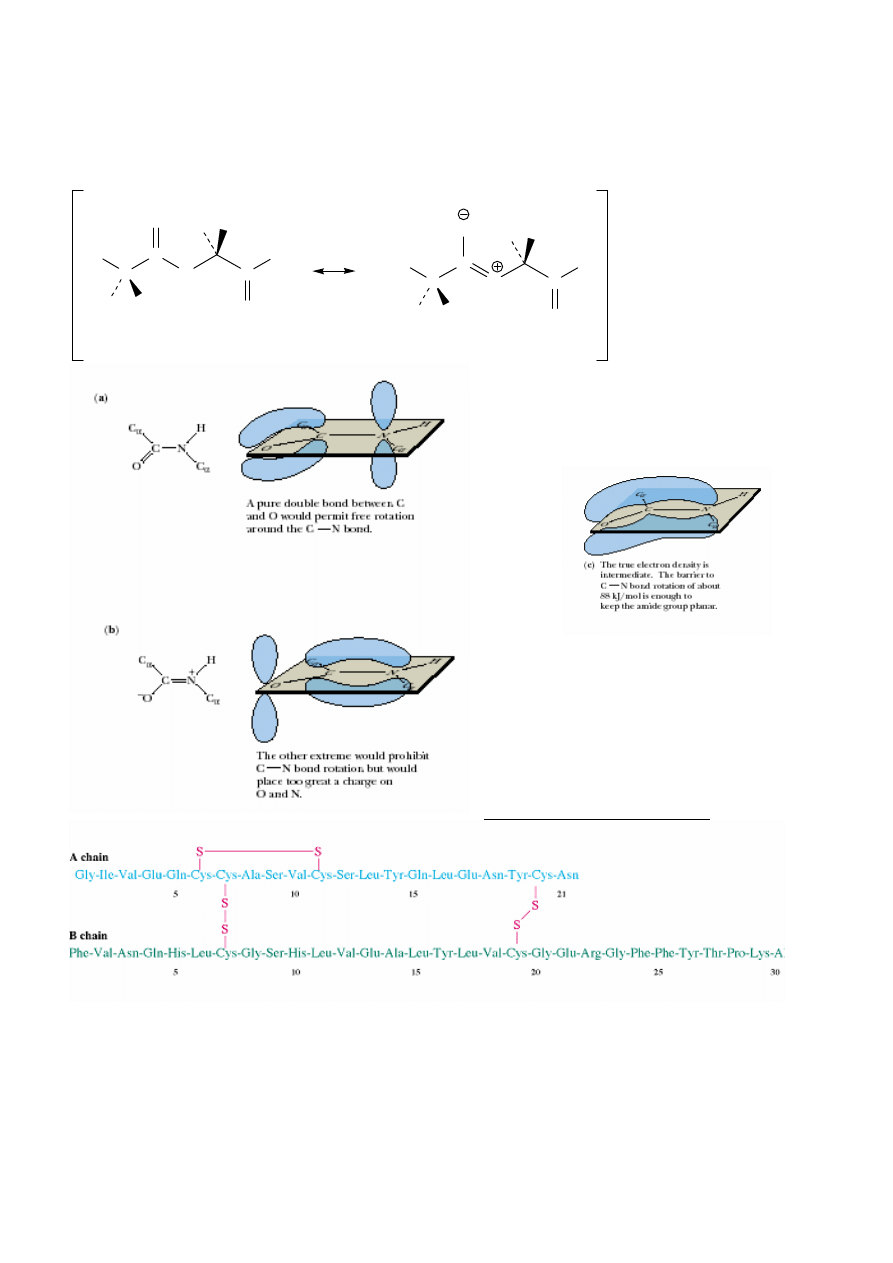
Sekwencja insuliny wołowej:
Kiedy
ś
– trzustka zabitych zwierz
ą
t; teraz – bakterie z genami ludzkiej insuliny.
Aspartam - Asp-Phe-OCH
3
Glutation -
γ
-Glu-Cys-Cys
Płasko
ść
wi
ą
zania amidowego, konfiguracja cis, oddziaływania mi
ę
dzy ła
ń
cuchami bocznymi
⇒
konfiguracja
trans
Minimalizowanie oddziaływa
ń
sterycznych i maksymalizowanie elektrostatycznych, dyspersyjnych
i wodorowych
⇒
⇒
⇒
⇒
12
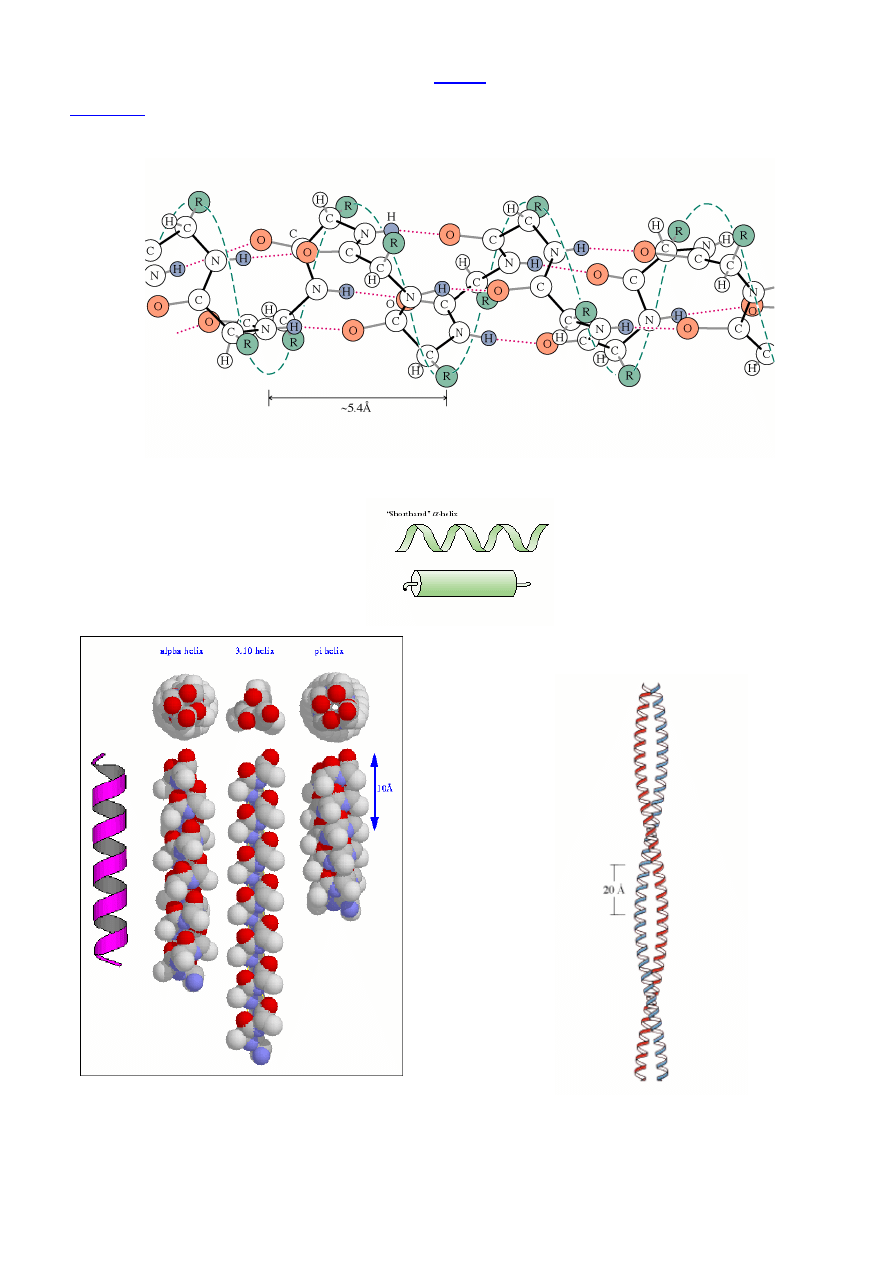
2° Struktura peptydu/białka –
lokalna konformacja ła
ń
cucha peptydowego
Charakterystyczne typy kształtu ła
ń
cucha polipeptydowego
wynikaj
ą
ce z oddziaływania niedaleko le
żą
cych reszt Aaa
←
X-ray, 2D-NMR
ββββ
-Kartki (harmonijka
β
, pofałdowany arkusz
β
,
β
-sheet) – równoległa i antyrównoległa
(np. fibroina jedwabiu)
Wi
ą
zania wodorowe mi
ę
dzy dwoma ła
ń
cuchami peptydowymi.
Równoległa
β
-kartka:
13
Helisy
αααα
-Helisa,
helisa
3,6
13
, (np. keratyna włosów) - wewn
ą
trzcz
ą
steczkowe w.wodorowe mi
ę
dzy C=O
reszty n i NH (n+4); s = 5.4
Ẩ
)
superhelisa
Lokalizacja reszt w globularnych białkach:

14
1. Reszty z niepolarnymi ła
ń
cuchami bocznymi (Val,Leu, Ile, Phe…) – wewn
ą
trz białka,
ograniczony kontakt z wodnym
ś
rodowiskiem;
2. Ła
ń
cuchy boczne reszt polarnych (Arg, Lys, Glu, Asp) – zwykle na powierzchni;
3. Nienaładowane polarne ła
ń
cuchy boczne (Ser, Thr, Trp, Tyr, Asn) – cz
ę
sto na powierzchni,
ale tak
ż
e, zwi
ą
zane wodorowo - wewn
ą
trz cz
ą
steczki.
Konformacja statystycznego kł
ę
bka.
3° Struktura peptydu/białka –
struktura trójwymiarowa wynikaj
ą
ca z dalszego fałdowania
ła
ń
cucha polipeptydowego.
Białka globularne (np. mioglobina, hemoglobina) – wi
ę
ksze fałdowanie (ekspozycja grup
hydrofilowych do
ś
rodowiska); odpowiedzialne za transport chemiczny i kataliz
ę
.
Białka fibrylarne
(np. miozyna – mi
ęś
nie, fibryna – skrzepy krwi,
α
-keratyna - włosy) –
„superhelisa”
Trzeciorz
ę
dowa struktura enzymów, białek transportuj
ą
cych – miejsce aktywne
Białka: fibrylarne, globularne, membranowe
4° Struktura białka -
sposób w jaki kilka ła
ń
cuchów polipeptydowych (podjednostki; domeny) ł
ą
czy
si
ę
w agregat.
Denaturacja – zniszczenie 3° struktury białka (
→
precypitacja, zniszczenie aktywno
ś
ci
katalitycznej…)
15
Okre
ś
lanie struktury 1° (sekwencjonowanie)
I.
Oczyszczanie polipeptydów:
1. Dializa;
2. Chromatografia s
ą
czenia molekularnego (gel-filtration chromatography);
3. Chromatografia jonowymienna;
4. Elektroforeza (> 1000 białek w 1 eksperymencie!);
5. Chromatografia powinowactwa (affinity chromatography).
II.
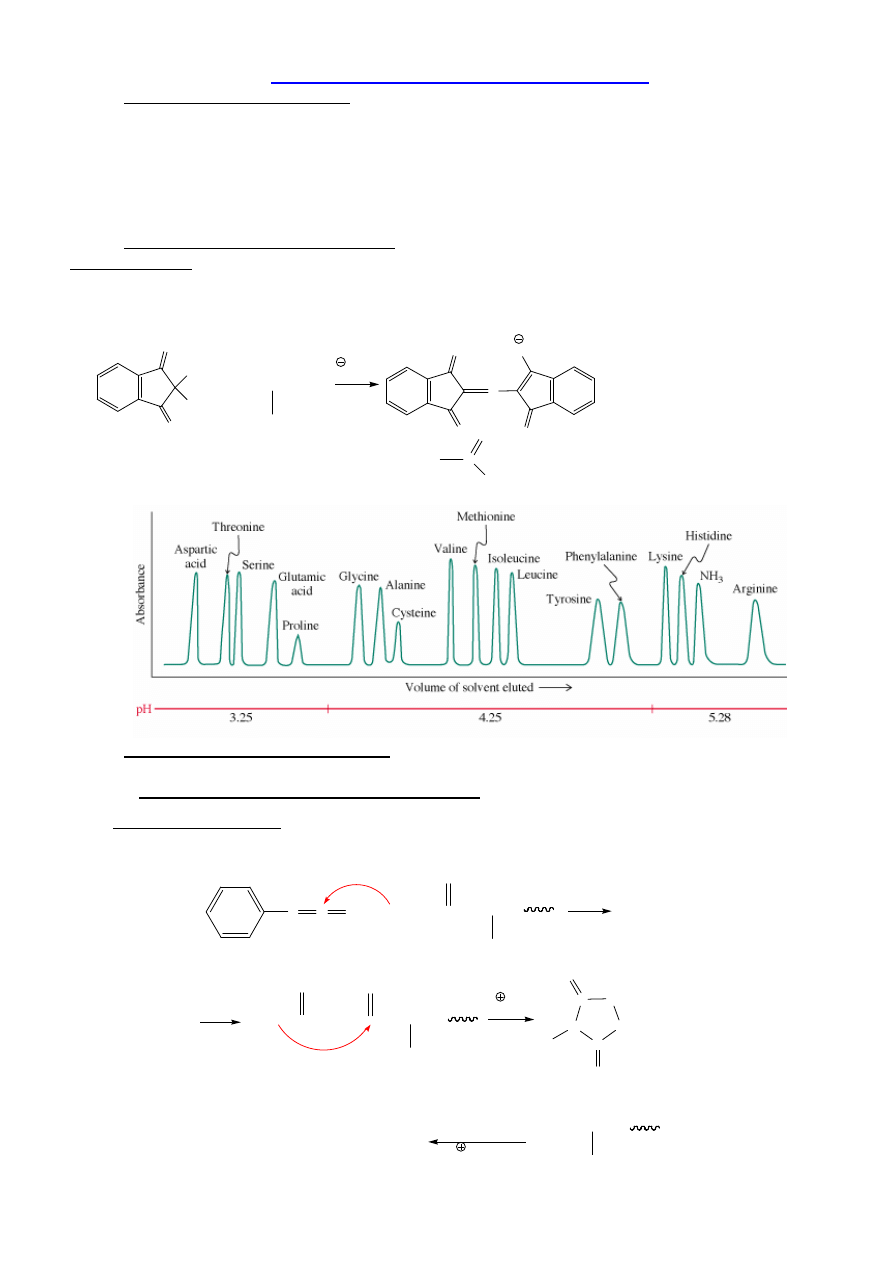
Badanie składu aminokwasowego: analizator aminokwasów lub MS
Analizator Aaa
a/ hydroliza (6 N HCl, 110 °C, 24 h);
b/ analiza aminokwasowa – kolumna z naładowanym ujemnie no
ś
nikiem, wymywanie
buforami, po elucji – reakcja z ninhydryn
ą
.
O
O
OH
OH
+ NH
2
CHCOOH
R
O
O
O
O
N
OH
H
2
O
+ R
C
O
H
+ CO
2
Absorbancja = f(t):
III.
Sekwencjonowanie tradycyjne:
a/ Ustalanie aminokwasu N-terminalnego:
Degradacja Edman’a:
Odczynnik Edmana = tiocyjanian fenylu
N
C
S
+ NH
2
-CH
2
-C-NH-CH-CO
O
CH(CH
3
)
2
Ph-NH-C-NH-CH
2
-C-NH-CH-CO
S
CH(CH
3
)
2
O
H, H
2
O
C
N
C
CH
2
NH
O
S
Ph
+
+ NH
2
-CH-CO
CH(CH
3
)
2
N-fenylotiohydantoina glicyny
PTH(Gly)
1. PhN=C=S
2. H, H
2
O
PTH(Val) + skrócony peptyd
16
Sekwenatory automatyczne - zwykle do 50 Aaa (nagromadzanie produktów ubocznych)
→
oznaczanie sekwencji w segmentach, inna hydroliza – oznaczenie uło
ż
enia segmentów.
Trypsyna – hydroliza przy COOH Lys, His
Chymotrypsyna – po Phe, Trp, Tyr
b/ Sekwencjonowanie z u
ż
yciem rekombinacyjnej technologii DNA
c/ Oznaczanie reszty C-ko
ń
cowej:
Inkubacja polipeptydu z karboksypeptydaz
ą
. Badania pojawiaj
ą
cego si
ę
Aaa. Peptyd z
nowym C-ko
ń
cem – dalsza degradacja.
Karboksypeptydaza A (trzustka wołowa) – prócz Pro, Arg, Lys
Karboksypeptydaza B (trzustka wieprzowa) – tylko gdy C-terminalna Arg lub Lys
Karboksypeptydaza A + B – wszystkie prócz Pro
Karboksypeptydaza C (li
ś
cie cytrusów) – wszystkie
Karboksypeptydaza Y (dro
ż
d
ż
e) – wszystkie
Strategia sekwencjonowania białek:
1. Gdy kilka ła
ń
cuchów polipeptydowych – rozdział (ekstremalne pH, 8M mocznik,
chlorowodorek guanidyny…).
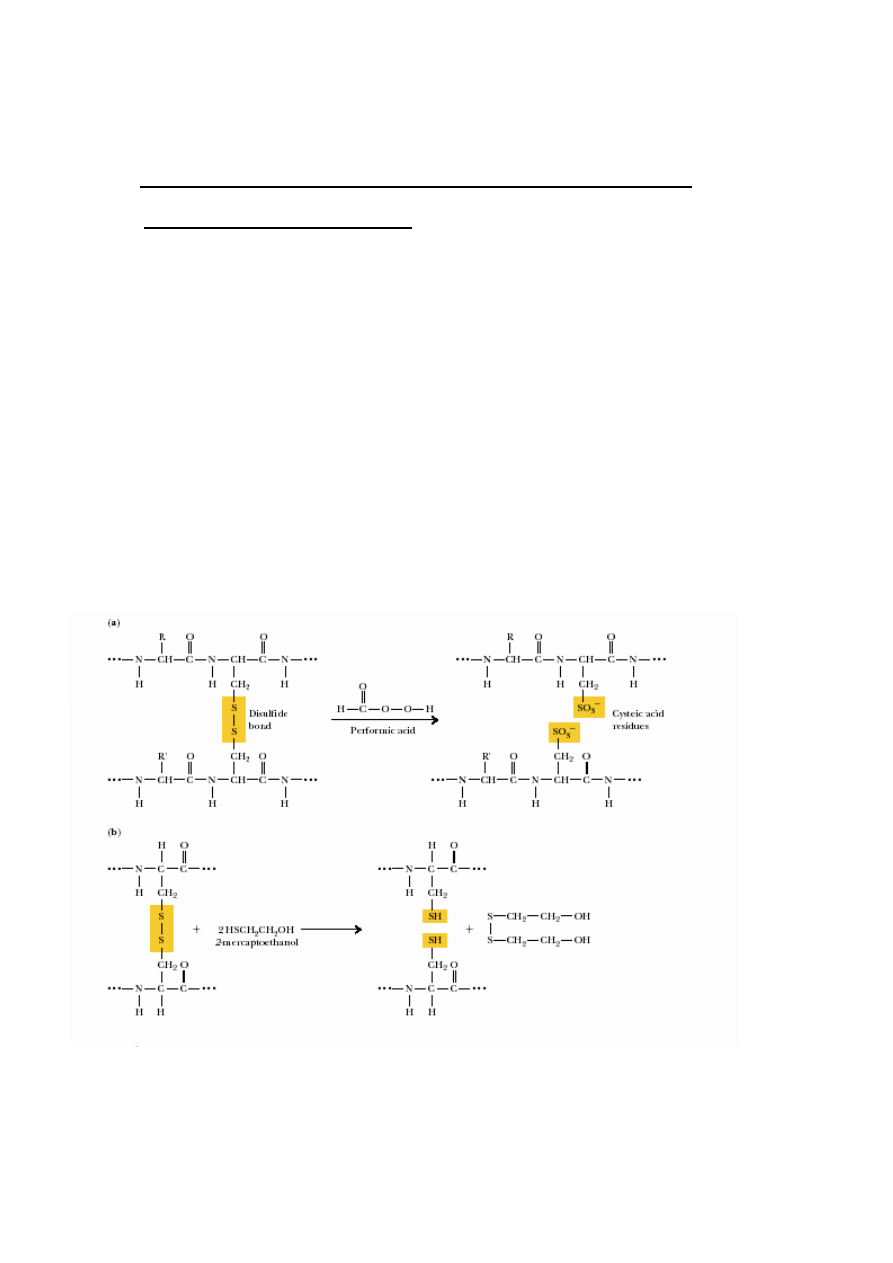
2. Rozerwanie mostków disiarczkowych:
Gdy S-S mi
ę
dzyła
ń
cuchowe 2
→
1
modyfikacje reaktywnego SH:
17
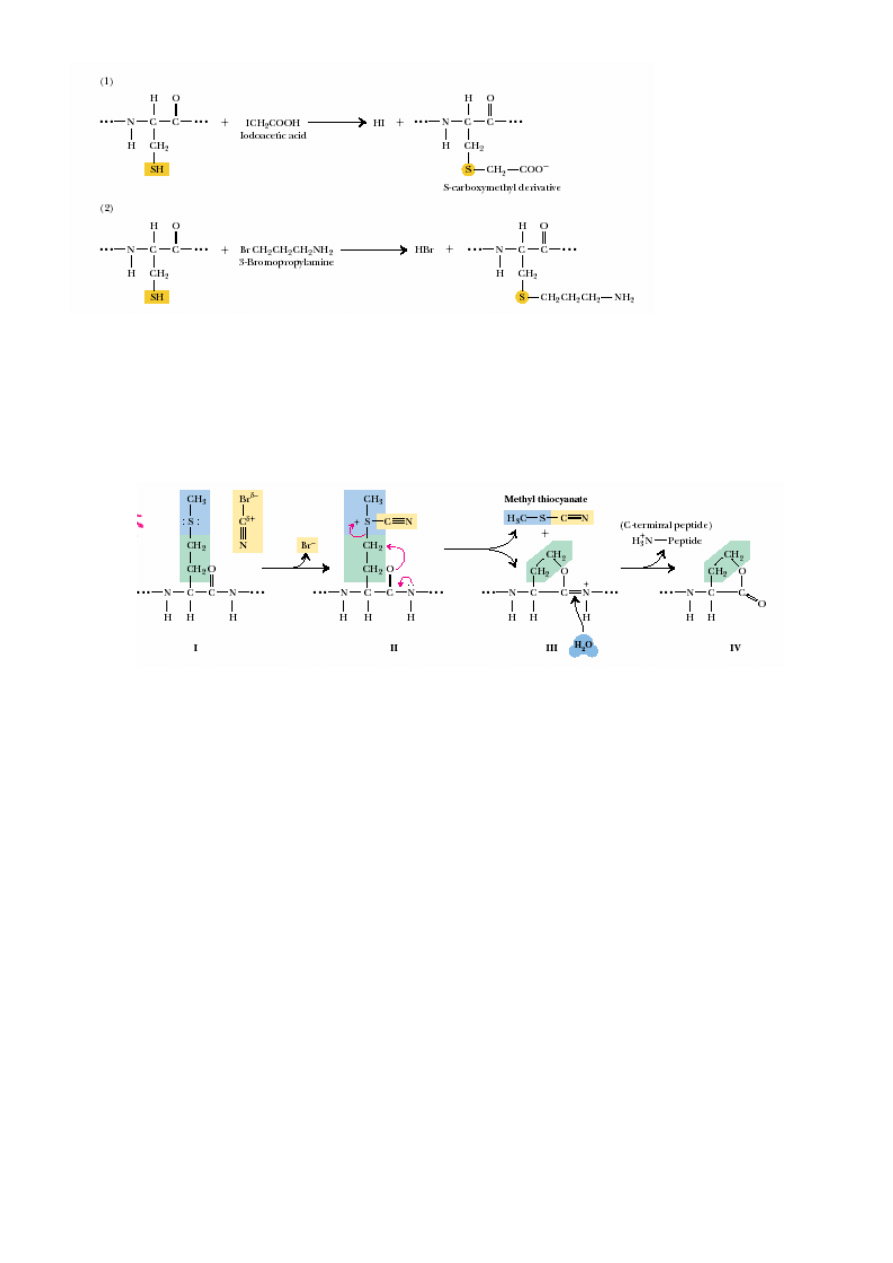
3. Ustalanie składu aminokwasowego.
4. Identyfikacja reszty C- i N-ko
ń
cowej (karboksypeptydazy, degradacja Edmana).
5. Hydroliza na mniejsze fragmenty, ustalanie ich sekwencji (trypsyna – po Arg, Lys,
chymotrypsyna – po aromatycznych Aaa, bromocyjan – po Met…).
lakton homoseryny
CNBr w 70% HCOOH
niestabilny w wodzie
produkt posredni
6. P-kt 5 powtórzony po innej proteolizie, generowanie nakładaj
ą
cych si
ę
fragmentów.
7. Rekonstruowanie sekwencji na podstawie nakładaj
ą
cych si
ę
fragmentów/
determinacja sekwencji na podstawie MS.
8. Lokalizacja mostków S-S.
Wyszukiwarka
Podobne podstrony:
01 Aminokwasy, peptydy, białka, enzymyid 3054 ppt
68 Aminokwasy peptydy i bialka(1)
68 Aminokwasy peptydy i bialka
Aminokwasy,peptydy,białka i inne
Aminokwasy, peptydy, białka
Chemia 6 Aminokwasy Peptydy i Białka
AMINOKWASY, PEPTYDY, BIAŁKA
Aminokw,peptydy,białka,enzymy
01 Aminokwasy, peptydy, białka, enzymyid 3054 ppt
Chemia 6 Aminokwasy Peptydy i Białka
Aminokwasy, peptydy, białka
Peptydy i białka
8 AMINOKWASY, PEPTYDY, BIA id 4 Nieznany (2)
Æwiczenie 7 Aminokwasy, peptydy i bia³ka
Peptydy i białka
AMINOKWASY I PEPTYDY
więcej podobnych podstron