
1. Trawienie pokarmów
Trawienie zaczyna się już w jamie ustnej, pokarm jest wstępnie rozkładany przez enzymy zawarte w ślinie
przy pH 7. Amylaza ślinowa rozkłada wielocukry na dwucukry, a skrobię na maltozę. Maltaza rozkłada
maltozę na glukozę, a Lizozym rozrywa wiązania glikozydowe.
Trawienie w żołądku odbywa się za udziałem soków żołądkowych przy pH 1,5. W soku znajdują się
enzymy takie jak; pepsyna, która skraca długoś łańcucha aminokwasów, renina, produkowana jest tylko u
młodych ssaków, Kazeina i parakazeina łączy się jonami wapnia i wytrąca z roztworu, Lipaza żołądkowa
działa na tłuszcze.
Trawienie w jelicie cienkim rozpoczyna się w dwunastnicy przy pH 8,5. Nie posiadają własnych enzymów,
ale pobiera je z wątroby i trzustki. W wątrobie produkowana jest żółć, która emulguje tłuszcze, rozbija je
w zawiesiny mikrokropelek, Trzustka produkuje trypsynę, chymotrypsynę, lipaze trzustkową oraz
amylazę trzustkową.
2. Rola gruczołów układu trawiennego
Wyróżniamy gruczoły przyścienne – mają mikskopijne wymiary, znajdują się w ścianach przewodu
pokarmowego. Zaliczamy tu gruczoły żołądkowe) wydzielają sok żołądkowy), gruczoły dwunastnicy
(wydzielają sok dwunastniczy), Gruczoły jelitowe (sok jelitowy). Mamy też gruczoły pozaścienne takie jak;
ślinianki w jamie ustnej, wątroba i trzustka. W skład soków trawiennych wchodzi woda, enzymy,
elektrolity oraz organiczne i nieorganiczne związki. W ciągu doby przewód wydziela ok.8-10L soku.
3. Enzymy trawienia białek, tłuszczów, węglowodanów i mechanizmy wchłaniania
Trawienie białek – białka są denaturowane przez soki żołądkowe, pepsyna rozczepia wiązania
peptydowe, Endopeptydazy (trypsyna, chemotrypsyna, elastaza) katalizuje rozpad specyficznych wiązań
peptydowych w jelicie, Egzopetydazy usuwają końce aminokwasów. Gdy „ścinają” N-końce są to
aminopeptydazy, a gdy C-koniec są to karboksypeptydazy, aminokwasy wchłaniają się przez błonę
śluzową jelita.
Trawienie tłuszczy – są emulowane przez sole kwasów żółciowych co ułatwia dalsze trawienie przez
enzymy. Odbywa się w jelicie cienkim głównie przez lipaze trójglicerydową. Lipazy są stymulowane
przez jony magnezu, kolipazę, fosfolipazę, sole kwasów żółciowych. Odcinają kolejne kwasy tłuszczowe
aż do powstania glicerolu oraz 2-monoacyloglicerolu. Chylomilrony odpowiadaj za transport
triglicerydów z jelita do mięśni i nerek, serca, mózgu itp. VLDL są przenośnikami triacyloglicerole
endogenne, LDL są przenośnikami cholesterolu i jego estrów. HDL – odpowiedzialny jest za transport
zwrotny cholesterolu z tkanki do wątroby.
Trawienie cukrów – rozpoczyna się w jamie ustnej, sacharydy rozkładają się przez amylazę ślinową do
oligosacharydów, które następnie są przez oligosacharydazy oraz disacharydazy rozkładane do
monocukrów, a one są wchłaniane do komórki na drodze transportu wtórnego aktywnego.
Produkty trawienia są wchłaniane przez błony komórkowe nabłonka jelitowego do krwi (aminokwasy,
cukry) do limfy (witaminy, produkty rozkładu tłuszczów), do jelita grubego (woda, sole żółciowe). W
jelicie cienkim występują kosmki, które zwiększają powierzchnię chłonną jelita do ok. 10m
2
. Kosmyki są
zaopatrzone w naczynia krwionośne oraz limfatyczne. Wchłanianie odbywa się przez pory w błonie
kosmka w procesie transportu aktywnego.
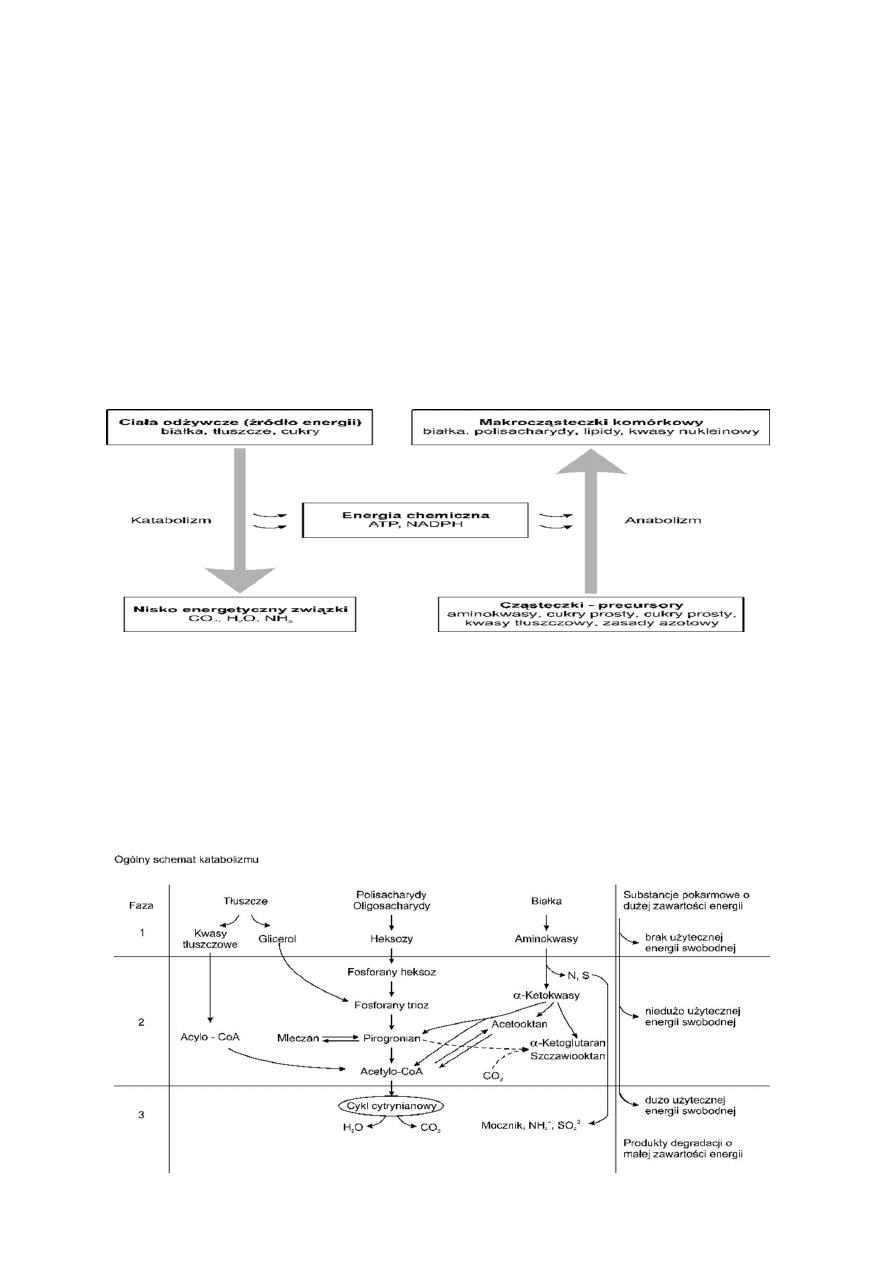
4. Ogólny schemat katabolizmu i anabolizmu
5. Fazy katabolizmu
Składa się z trzech faz; Faza 1- Substancje takie jak Tłuszcze białka, polisacharydy są trawione w układzie
pokarmowym do mniejszych cząsteczek. Faza 2- Wcześniej rozdrobnione cząsteczki są transportowane i
trawione go jeszcze mniejszych cząsteczek, głównie Acetylko-CoA podczas tych przemian uwalniana jest
niewielka ilość energii swobodnej. Faza 3- Acetylo-CoA jest utleniany w cyklu Krebsa, podczas tego
procesu powstaje duża ilość energii swobodnej. Produktami końcowymi jest CO
2
i H
2
O.

6. Związki makroergiczne
Guanozynotrifoforan (GTP) – dostarcza energię do procesów syntezy białek . Jest również
modulatorem struktury przestrzennej białka. Cytrydynotrifosforan (CTP) – jest dawcą energii
w procesy syntezy lipidów. Urydynotrifosforan (UTP) – jest wykorzystywany w procesach
syntezy polisacharydów. fosfofenylopirogronian (PEP)
Kreatynina – kwas β-metyloguanidynooctowy – Kreatyna jest stałym składnikiem tkanki
mięśniowej i odgrywa zasadniczą rolę w przemianach chemicznych powodujących skurcz
mięśniowy. Fosfokratynina wpływa stymulująco na anabolizm.
Acetylko-CoA – acetylokoenzym A jest tioestrem, służy jako przenośnik grup acetylowych.
7.
Glikoliza i glikogeneza,
przebieg i zys energetyczny
Glikoliza
Glikogeneza
Jest to szereg reakcji prowadzących do przekształcenia
glukozy w pirogronian.
Jest to proces syntezy glikogenu,
cząsteczki glukozy są dodawane do
kolejnych
łańcuchów
glikogenu.
Wpływają na ten proces hormony takie
jak;
adrenalina,
noradrenalina,
insulina.
FAZA I – PRZEKSZTAŁCENIE GLUKOZY W 1,6-
BISFOSFORAN
Reakcja 1 – Fosforylacja glukozy przy udziale
heksozokinaz. ATP ADP
Glukoza ⇔ Glukozo-6-
Fosforan
Reakcja 2 – izomeryzacja przy udziale izomerazy
glukozofosforanowej.
Glukozo-6-fosforan
Fruktozo-6-Fosforan
Reakcja 3 – Fosforylacja przy udziale
fosfofruktokinazy.
Fruktozo-6-Fosforan 1,6-
Bisfosforan
FAZA II – PRZEKSZTAŁCENIE 1,6-BISFOSFORANU W
PIROGRONIAN
Proces ten przebiega tak samo jak
glikoliza, jedynie trzy reakcje, które nie
są odwracalne są inne.
Reakcja 1- Glukoza jest przyłączana się
z udziałem ATP i przemienia się pod
wpływem heksokinay.
Glukoza Glukozo-6- fosforan
Reakcja 2- W przemianie uczestniczy
fosfoglukomutaza.
Glukozo-6- fosforan Glukozo-1-
Fosforan
Reakcja 3 – w przmeianie uczestniczy

Reakcja 4- Zachodzi rozczepienie 1,6-bisfosforan przy
udziale aldolazy.
Fruktozo-1,6-bisfosforan Fosforan
dihydroksyacetonu, Gliceroaldehyd 3-fosforan
Reakcja 5- Izoreryzacja przy udziale izomerazy
trizofosforanowej.
Fosfodihydroksyaceton Aldehyd
3-fosfogliceryacetonowy (GAP)
Reakcja 6- Utlenienie przy udziale dehydrogenazy
aledehydu fosfoglicerynowego.
GAP ⇔ 1,3-
bisfosfoglicerynian
Reakcja 7- defosforylacja katalizowana przez kinazy
fosfoglicerynowe.
1,3-bisfosfoglicerynian ⇔ 3-
fosfoglicerynian
Reakcja 8- Przegrupowanie wewnątrzcząsteczkowe
przy udziale fosfogliceromutazy.
3-fosfoglicerynian
⇔ 2-fosfoglicerynian
Reakcja 9- Przemiana przy udziale enolaz.
2-
fosfoglicerynian ⇔ Fosfoenolopirogronian
Reakcja 10 – Tworzenie pirogronianu przy udziale kinaz
pirogronianowych.
Fosfoenolopirogronian
Pirogronian
pirofosforylaza UDP- glukozy
Glukozo-1-Fosforan UDP-Glukoza
Reakcja
4-
Zachodzi
synteza
glikogenowa zachodzi przeniesienie
glukozy z UDP-glukozy do rosnącego
łańcucha polisacharydowego. Reszty
glukozy dołanczane są do grupy- OH,
tworząc wiązanie α-1,4-glikozydowe.
2 ATP
8. Rola hormonów w regulację procesu
Insulina – aktywuje syntezę glikogenową i kilka enzymów glikolizy. Insulina jest wydzielana
przez trzustkę, bierze udział w metabolizmie białek, węglowodanów i tłuszczów. Cząsteczka
insuliny łączy się z receptorem insulinowym. Napływ insuliny do krwi odbywa się za
pośrednictwem specjalnych przenośników GLUT. Receptor insulinowy składa się z α-β-β-α jest
to heterotetramer połączony wiązaniami siarkowymi. Receptory licznie występują na
powierzchni komórek wątroby, mięśni i adipocytów.
Glukagon i adrenalina – indukują enzymy glikogenazy i hamują działanie kinazy
pirogronianowej. Wpływa na syntezę cAMP – hamuje syntezę i aktywuje rozkład glikogenu.

Kortyzol – indukuje enyzmy glukonogenezy i rozkład aminokwasów – prekursorów
glukonogenezy
Enzymy amylolityczne – rozkładają skrobię, glikogen i disacharydy w przewodzie
pokarmowym. Zaliczamy tu enzymy należące do hydrolaz zaliczamy tu α-amylazę (rozkłada
wiązania α-1,4-glikozydowe wewnątrz łańcucha polisacharydowego, początkowo tworzą
dekstryny, które potem maltozy lub izomaltozy) , α-1,6-glikozydaza (rozkłada wyłącznie
wiązania α-1,6-glikozydowe) , maltoza, laktoza, sacharaza, glukoamylaza (rozkłada wiążania
α-1,4-glikozydowe od nieredukującego końca skrobi, jako produkt daje cząstkę glukozy i
dekstrynę).
Β-amylaza – występuje na nasionach różnych roślin i rozkłada skrobię w trakcie kiełkowania.
Hydrolizują co drugie wiązanie α-1,4-glikozydowe produktami jest dekstryna i β-maltoza.
9.
Los pirogranianu i cykl Corich
Tworzenie pirogronianu jest katalizowane przez kinaza pirogronianowa. Glikoliza - oddychanie
tlenowe zachodzi w mitochondrium wyróżniamy tworzenie acetylo-CoA, które wchodzi do
cyklu Krebsa, a następnie transport elektronów i chemiosmoza (pirogronian acetylo-CoA
dalsze utleniane). W oddychaniu beztlenowym pirogronian po glikolizie ulega fermentacji
alkoholowej(pirogronian aldehyd octowy etanol) i fermentacji mleczanowej (
pirogronian mleczan).
Fementacja alkoholowa – pirogronian ulega dekarboksylacji do aldehydu, który jest
redukowany do etanolu przy utlenieniu NADH.
Fermentacja mlekowa- pirogranian jest redukowany d mleczanu przy jednoczesnym
utlenieniu NADH.
Cykl Corich
Inaczej jest to cykl kwasu mlekowego, jest to ciąg przemian,w których mleczan powstaje w
beztlenowej glikolizie w mięśniach i erytrocytach, jest transportowany przez krew do
wątroby. Tam jest przetwarzany w procesie glukonogenezy.
10. Ogólny schemat szlaku pentozo-fosforanowego i fazy tego procesu
Szlak pentozofosforanowy ciąg reakcji biochemicznych, podczas których glukozo-6-fosforan
jest utleniany do rybulozo-5-fosforanu oraz wytwarzany jest NADPH. Reakcje szlaku zachodzą w

cytozolu. Przede wszystkim w tkance tłuszczowej, gruczołach mlecznych i korze nadnerczy oraz
cytoplazmie i plastydach komórek roślinnych.
Faza 1 – oksydacyjna.
Reakcja 1; Uczestniczą tu dehydrogenaza glukozo-6-fosforanowa.
glukozo-6-fosforan +
2NADP
+
6-fosfoglukono-δ-lakton+ NADPH+H+
Reakcja 2 – Uczestniczy tu 6-glukonolaktonaza.
6-fosfoglukono-δ-lakton 6-fosfoglukonian +
H+
Reakcja 3 – Uczestniczy tu dehydrogenaza 6-fosfoglukonianowa.
6-fosfoglukonian
rybulozo-5-fosforan + NADPH
+
+ H
+
+ CO
2
Faza 2 – nieoksydacyjna
Reakcja 1 –Uczestniczy w tym izomeraza pentozofosforanowa .
rybulozo-5-fosforan
rybozo-5-fosforan
Reakcja 2-Uczestniczy w tym epimeraza pentozofosforanowa.
rybozo-5-fosforan
ksylulozo-5-fosforan
Reakcja 3- Uczestniczy w tym transketolaza.
ksylulozo-5-fosforan+ rybozo-5-fosforan
aldehyd 3-fosfoglicerynowy + sedoheptulozo-7-fosforan
Reakcja 4- Uczestniczy w tym transketolaza.
sedoheptulozo-7-fosforan + aldehyd 3-
fosfoglicerynowy erytrozo-4-fosforan + fruktozo-6-fosforan
Reakcja 5- Uczestniczy w tym transketolaza.
ksylulozo-5-fosforan + erytrozo-4-fosforan
aldehyd 3-fosfoglicerynowy + fruktozo-6-fosforan
11. Ogólna charakterystyka procesów oksydacji.
-oksydacja i biosynteza kwasów
tłuszczowych nasyconych.
α-oksydacja - polega na 2 hydroksylacji z udziałem oksygenazy mieszanej, odwodornieniu z
udziałem dehydrogenazy zależniej od NAD
+
i dekarboksylacji oksydacyjnej powstałego 2-
oksokwasu dając kwas tłuszczowy o łańcuchu skróconym o 1 atom węgla.
β-oksydacja- jest procesem dostarczającym równoważników redukcyjnych służących w
łańcuchu oddechowym wytworzeniu ATP, acetylo-CoA do cyklu Krebsa służącemu wytworzeniu

ATP, w wątrobie substratów do syntezy ciał ketonowych, zwłaszcza w przypadku zaburzeń
gospodarki cukrami. U eukariotów zachodzi w macierzy mitochondrialnej, a u prokariotów w
cytoplazmie. Reakcje β-oksydacji polegają na takich przemianach by rozczepić "dłuższy" acylo-
CoA na acetylo-CoA i acylo-CoA "krótszy", po czym rozpocząć proces od początku, aż do
momentu gdy powstają dwie cząteczki acetylo-CoA w przypadku kwasów tłuszczowych o
parzystej liczbie węgli lub propionylo-CoA i acetylo-CoA w przypadku kwasów o nieparzystej
liczbie węgli.
Reakcja 1 – Utlenienie, za pomocą dehydrogenazy acylo-CoA,
acylo-CoA
trans-Δ2-enoilo-
CoA
z wytworzeniem FADH2.
Reakcja 2- Uwodnienie
trans-Δ2-enoilo-CoA
3-hydroksyacylo-CoA
za pomocą enzymu
hydrataza enoilo-CoA.
Reakcja 3- Utlenienie
3-hydroksyacylo-CoA
3-ketoacylo-CoA
za pomocą dehydrogenazy
hydroksyacylo-CoA i z wytworzeniem NADH.
Reakcja 4-
Tioliza 3-ketoacylo-CoA
przez drugą cząsteczkę CoA i wytworzenie
acylo-CoA
skróconego o dwa atomy węgla oraz
acetylo-CoA
. Katalizatorem w tej reakcji jest β-ketotiolaza.
ω- oksydacja – w jej wyniku powstają krótkie kwasy dikarboksylowe, zachodzi w retikulum
endoplazmatycznym. est katalizowany przez monooksygenazęcytochromu P-450 używającą tlen
O
2
jako substratu.
Biosynteza nasyconych kwasów tłuszczowych - reakcje biochemiczne, prowadzące do
powstania kwasów tłuszczowych z jednostek acetylo-CoA, zachodzi w cytoplazmie
Reakcja 1- syntezy jest karboksylacja acetylo-CoA do malonylo-CoA, katalizowana przez
karboksylazę acetylo-CoA. Następnie acetylo-CoA łączy się z białkowym nośnikiem grup
acylowych (ACP), w wyniku czego powstaje acetylo-ACP, a z malonylo-CoA powstaje malonylo-
ACP.
Reakcja 2 - są cykle elongacji, które można podzielić na 4 fazy (kondensacja, redukcja,
odwodnienie, redukcja).
Faza 1- Kondensacja 2-węglowego acetylo-ACP i 3-węglowego malonylo-ACP do 4-węglowego
acetoacetylo-ACP. Podczas tej reakcji zostaje odłączony jeden ACP oraz CO2. Enzym katalizujący
ten etap to enzym kondensujący acylomalonylo-ACP

Faza 2 - Redukcja acetoacetylo-ACP do D-3-hydroksybutyrylo-ACP, podczas której
wykorzystywana jest jedna cząsteczka NADPH. Enzym - reduktaza beta-ketoacylo-ACP
Faza 3- Odwodnienie D-3-hydroksybutyrylo-ACP do krotonylo-ACP. Enzym- dehydrataza 3-
hydroksyacylo-ACP
Faza 4 - Redukcja krotonylo-ACP do butyrylo-ACP, podczas której zostaje wykorzystana kolejna
cząsteczka NADPH. Enzym: reduktaza enoilo-ACP
12. Degradacja białka
Proces ten może zachodzić trzema drogami: przez układ lizosomalny, ubikwitynowy
(proteasomowy) oraz cytoplazmatyczny (kalpainy).
Degradacja w lizosomach – lizosomy odpowiadają za degradację białek, maja pH 5,0. Posiadają w
sobie enzymy z grupy hydrolaz umożliwiające degradację. Degradacja białek przebiega
wieloetapowo, przez stopniowy rozpad struktury białkowej. Degradacji ulegają białka
egzogenne i „długowieczne” białka endogenne, np. białka strukturalne.
Proteasom rozkłada jedynie białka, które uległy „wyznakowaniu” w procesie ubikwitynacji.
Proces ten polega na przyłączeniu do cząsteczki białka mającego ulec destrukcji, cząsteczki
małego białka – ubikwityny, będącym swoistym znacznikiem molekularnym. Białka
„wyznakowane” cząsteczką ubikwityny kierowane są bezposrednio do proteasomów. Taki
mechanizm umożliwia wybiórczy rozkład białek, które nie są już potrzebne. W proteasomach
degradowane są białka krótkożyjace, np. enzymy, zawierające na N-końcu określoną sekwencję
aminokwasów, rozpoznawalną przez ubikwitynę lub białka, które zostały wadliwie
zsyntetyzowane.
Kalpainy są nielizosomowymi proteazami cysteinowymi występującymi w cytoplazmie
komórek. Aktywność proteolityczna kalpain jest zależna od poziomu jonów Ca2+. Degradują one
białka nieselektywnie. Obserwuje się raczej powinowactwo do trzeciorzędowej struktury białka
niż jego sekwencji aminokwasowej (np. sekwencji sygnałowej na N-końcu) oraz do białek i
peptydów zawierających aminokwasy hydrofobowe (leucyna, walina i izoleucyna) i
aromatyczne (fenyloalanina i tyrozyna).
13. Proteosoma i rola ubikwityny
Proteasom jest kompleksem białkowym odpowiadającym za degradację białek. Występują w
jądrze i cytoplazmie. Zdegradowane białka są oznakowane za pomocą małych białek tzw.

Ubikwityna. Jedna cząsteczka tego białka jest przyłączana za pomocą ligazy ubikwitynowej, co
umożliwia identyfikacje białka przeznaczonego do degradacji. Proteasom ma kształt beczki,
składa się z czterech pierścieni ułożonych wokół centralnego porów. Każdy z tych pierścieni jest
złożony z siedmiu pojedynczych białek. Dwa pierścienie wewnętrzne zawierają β podjednostek
białkowych, tworzących miejsca aktywne. Dwie zewnętrzne pierścienie zawierają α
podjednostkę, którego funkcją jest utrzymanie "drzwi", przez które można wprowadzić do
białek lufy. Proteoliza komórkowa pełni wiele funkcji: uaktywnia bądź unieczynnia enzymy,
moduluje wewnątrzkomórkowe przekazywanie sygnałów, usuwa nadmiar produktów syntezy, i
co najważniejsze, rozkłada egzogenne i niepotrzebne endogenne, a także nieprawidłowo
zsyntetyzowane białka i peptydy
Ubikwityna – to małe białko, służy do degradacji białek jako znacznik, które maja ulec
proteolizie. Ubikwityna jest peptydem złożonym z 76 reszt aminokwasowych. Potranslacyjna
modyfikacja białek w procesie ubikwitynacji polega na przyłączaniu do reszt lizynowych danego
białka grupy karboksylowej C-końca reszty .
14.
Rozkład aminokwasów - procesy deaminacji, dekarboksylacji, rodziny aminokwasów.
Deaminacja - inaczej nazywana dezaminacją, polega na eliminacji grupy aminowej –NH
2
, z
wydzieleniem amoniaku. Grupa aminowa często jest przekształcana do grupy ketonowej. Jest to
pierwszy proces ich degradacji co umożłiwia wykorzystanie tych związków potem jako
substratu oddechowego. W naturalnym środowisku nie wymaga obecności tlenu ale warunkiem
jest odpowiednia ilość azotanów i organizmów nitryfikujących. Przeprowadzana jest przez
bakterie E. coli wytwarzające enzymy ureazy i Deaminazy.
Dekarboksylacja – jest to reakcja podczas, której dochodzi do usunięcia grupy karboksylowej z
kwasów karboksylowych, ich soli lub estrów. α-dekarboksylacja – jest to odrywanie grupy
karboksylowej przy α-atomie węgla, ω-dekarboksylacja charakterystyczna tylko dla
mikroorganizmów.
Rodziny aminokwasów;
Aminokwasy powstające z α-ketoglutaranu, który pochodzi z cyklu kwasu cytrynowego. Do
tej grupy należy tzw. rodzina glutaminianu:
glutamina, prolina i arginina
.
Aminokwasy powstające ze szczawiooctanu, który pochodzi z cyklu kwasu cytrynowego. Do
tej grupy należy tzw. rodzina asparaginianu:
asparagina, metionina, treonina
i
lizyna
oraz
izoleucyna
.

Aminokwasy powstające z 3-fosfoglicerynianu, który pochodzi z glikolizy. Do tej grupy
należy tzw. rodzina seryny;
seryna, cysteina i glicyna
.
Aminokwasy powstające z pirogronianu, który pochodzi z glikolizy. Do tej grupy należy
tzw. rodzina pirogronianu;
alanina, walina i leucyna
.
Aminokwasy powstające z fosfoenolopirogronianu, pochodzącego z glikolizy i erytrozo-4-
fosforanu, pochodzącego ze szlaku pentozofosforanowego. Do tej grupy należy tzw. rodzina
aminokwasów aromatycznych;
fenyloalanina, tyrozyna i tryptofan
.
Aminokwas
powstający
z
rybozo-5-fosforanu,
pochodzącego
ze
szlaku
pentozofosforanowego;
histydyna
.
15. Cykl Krebsa i jego rola.
Ma za zadanie dostarczenie równoważników redukujących zamienianych na energię
magazynowaną w ATP w łańcuchu oddechowym, dostarczać ma także energie w postaci GTP,
oraz dostarczać ważnych prekursorów do syntezy innych cząsteczek. Jest to końcowy wspólny
szlakiem utleniania cząsteczek będących źródłem energii dla organizmu, takich jak białka, kwasy
tłuszczowe, węglowodany.
Głównym substratem cyklu jest Acetylo-CoA, powstaje z pirogronianu w reakcji katalizowanej
przez kompleks dehydrogenazy pirogronianowej w mitochondriom.
Pirogronian Acetylo-
CoA.
Drugim substratem jest szczaiooctan odnawiany w tym cyklu, pod wpływem karboksylazy
pirogronianowej.
Pirogronian szczawiooctan
Reakcja 1 –Jest to proces kondensacji, który katalizowany jest przez syntezę cytrynianu.
Inhibitorami tego etapu są ATP.
Acetylo-CoA + szczawiooctan + H
2
O Cytrynylo-CoA wolny
koenzym A + cytrynian
Rakcja 2 – Dehydratacja (odwodnienie, uwodnienie) uczestniczy w tym enzym akonitaza.
Cytrynian cis-Akonityna + H
2
O izocytrynian
Reakcja 3- Utlenianie oraz dekarboksylacja uczestniczy w tym enzym dehydrogenaza
izocytrynianowa.
Izocytrynian + NAD
+
szczawiobursztynian +NADH
+
+H
+
α-ketoglutaran +
CO
2
.

Reakcja 4- Zachodzi dekarboksylacja i utlenianie przy udziale kompleksu dehydrogenazy α-
ketoglutaranowej.
α-ketoglutaran + NAD + CoA Bursztynylo-CoA +CO
2
+ NADH
.
Reakcja 5- Zachodzi fosforylacja substratowa przy udziale Tiokinazy bursztynianowej.
Bursztynylo-CoA +Pi +GAD Bursztynian +GTP +CO
2
.
Reakcja 6 – Utlenianie przy udziale dehydrogenazy bursztynianowej.
Bursztynian + FAD
Fumaran + FADH
2
.
Reakcja 7- Zachodzi addycja przy udziale fumarazy.
Fumaran + H
2
O Jabłczan
Reakcja 8 – Utlenianie przy udziale dehydrogenazy jabłczanowej.
Jabłczan + NAD
+
Szczawiooctan + NADH + H
+
.
16. Cykl mocznikowy.
Mocznik tworzy się z amoniaku i dwutlenku węgla oraz asparaginianu. Grupy aminowe
mocznika pochodzą z asparaginianu. Reakcje cyklu zachodzą w wątrobie, przy czym synteza
karbamoilofosforanu i cytruliny odbywa się w mitochondrium a synteza arginylobursztynianu i
uwalnianie mocznika odbywa się w cytozolu.
Bilans energetyczny syntezy mocznika: proces kosztowny energetycznie, ponieważ wytworzenie
jednej cząsteczki mocznika wymaga wkładu energetycznego 3 moli ATP: 2 mole do syntezy
karbamoilofosforanu (2 cząsteczki ATP przekształcają się do 2 cząsteczek ADP) i mol do syntezy
arginilobursztynianu (ATP przekształca się w AMP). Cykl jest niezbędny w celu usuwania
toksycznego amoniaku.
Reakcja 1 -
Ornityna
najpierw jest karbamoilowana przez karbamoilofosforan, co daje
cytrulinę
.
Reakcja 2 -
Cytrulina
ulega kondensacji z asparaginianem, tworząc argininobursztynian, który
jest rozszczepiany na
argininę i fumaran
.
Reakcja3 - Bezpośrednim prekursorem mocznika jest
arginina
, która z udziałem arginazy ulega
hydrolizie do
mocznika i ornityny
.
Reakcja 5. - Najpierw na
ornitynę
zostaje przeniesiona grupa karbamoilowa, co prowadzi do
otrzymania
cytruliny
. Reakcje te katalizuje karbamoilotransferaza ornitynowa. Donorem
karbamoilu w tej reakcji jest karbamoilofosforan, który dzięki wiązaniu bezwodnikowemu
charakteryzuje się dużym potencjałem przenoszenia.

Karbamoilofosforan niezbędny do utworzenia cytruliny jest syntetyzowany z NH4+, CO2, ATP i
H2O w skomplikowanej reakcji katalizowanej przez syntazę karbamoilofosforanową.
Reakcja 6 - W następnym etapie syntetaza
argininobursztynianowa
katalizuje kondensację
cytruliny z asparaginianem do
argininobursztynianu
. Reakcja ta przebiega kosztem energii ATP,
który ulega przy tym hydrolizie do AMP i pirofosforanu, oraz dzięki następnej hydrolizie
pirofosforanu.
Reakcja 8 - W końcu ligaza argininobursztynianowa rozszczepia
argininobursztynian
na
argininę i fumara
n. Szkielet węglowy asparaginianu nie zostaje podczas tej reakcji naruszony,
natomiast grupa aminowa zostaje przeniesiona na powstającą argininę.
Istotne znaczenie w cyklu mocznikowym ma synteza fumaranu, przez którą cykl mocznikowy
zazębia się z cyklem kwasu cytrynowego.
17. Transport elektronów w łańcuchu oddechowym i fosforylacja oksydacyjna
.
Transport elektronów ;
Oksydoreduktaza NADH-koenzym Q - jest pierwszym białkiem łańcucha transportu
elektronów. Reakcja katalizowana przez enzym polega na przeniesieniu dwóch elektronów z
NADH na koenzym Q10.
Oksydoreduktaza bursztynian-ubichinon - jest drugim punktem wejścia elektronów do
łańcucha transportu elektronów.
Oksydoreduktaza flawoproteina przenosząca elektron-ubichinon - jest trzecim punktem
wejścia do łańcucha transportu elektronów
Oksydoreduktaza koenzym Q-cytochrom c - Cytochromy są białkami przenoszącymi
elektrony, zawierającymi jedną lub więcej grup hemowych.
Oksydaza cytochromu c - jest ostatnim enzymem łańcucha transportu elektronów. Enzym
katalizuje końcową reakcje łańcucha oddechowego, przenosząc elektrony na tlen i jednocześnie
przemieszczając protony przez błonę
Fosforylacja oksydacyjna - jest szlakiem metabolicznym, w którego wyniku energia uwalniana
podczas utleniania zredukowanych nukleotydów przekształcana jest w energię ATP. w wyniku
szeregu reakcji redoks, elektrony przenoszone są ze zredukowanych nukleotydów, NADH i
FADH2, na pełniący funkcję akceptora elektronów tlen. Reakcji redoks zachodzi na kompleksach
białkowych znajdujących się w mitochondriach. Zestaw enzymów biorących udział w
przenoszeniu elektronów określa się jako łańcuch oddechowy. Przepływ elektronów z donorów
w postaci cząsteczek NADH na akceptory w postaci cząsteczek tlenu, odbywający się przez
szereg przenośników biorących udział w łańcuchu transportu elektronów, jest procesem
egzoenergetycznym – uwalniającym energię. Synteza ATP jest zaś procesem
endoenergetycznym który, aby zachodzić, wymaga dostarczenia energii. łańcuch oddechowy, jak
i synteza ATP zachodzi na błonach białkowo-lipidowych. Energia z łańcucha transportu
elektronów jest przenoszona na syntazę ATP, dzięki wytworzeniu różnicy stężeń jonów w

poprzek błony, nazywanej gradientem elektrochemicznym. Proces przenoszenia protonów
przez błonę został nazwany chemiozmozą Protony są przenoszone z apomocą pompy
protonowej. Powstający, w efekcie ich pracy, gradient elektrochemiczny, nazywany często siłą
protonomotoryczną, składa się z dwóch elementów: różnicy stężeń protonów raz różnicy
potencjałów, wynikającej z ładunków przemieszczanych cząsteczek. Syntaza ATP zużywa
energię gradientu elektrochemicznego, pozwalając przejść ładunkom (protonom) z powrotem
przez błonę.
Wyszukiwarka
Podobne podstrony:
Zagadnienia z BIOCHEMII-oprac studentow z korekta KM, Biotechnologia POLSL, Semestr V, Biochemia, Eg
BIOCHEMIA EGZAMIN - opracowane zagadnienia, Wychowanie Fizyczne (materiały i notatki)), Biochemia, B
Biochem, Akademia Wychowania Fizycznego, Biochemia, egzaminy
Biochemia, (Sylwia) studia semestr 3, Biochemia, EGZAMIN, EGZAMIN, egzam
Biochemia egzamin
Biochemia 2006 2007, materiały medycyna SUM, biochemia, EGZAMIN, teoretyczny
Biochemia egzamin[1]
Żelbet - Egzamin - Zagadnienia, Budownictwo S1, Semestr IV, Konstrukcje betonowe, Egzamin
BIOCHEMIA EGZAMIN
egzamin zagadnienia wf
Biochemia egzamin
na egzamin zagadnienia
Biochemia egzamin sciaga, BIOCHEMIA
Pedagogika społeczna, Egzamin - zagadnienia, Pedagogika społeczna-pytania i odpowiedzi na egzamin
pytania biochemia egzamin 12 I termin (1)
więcej podobnych podstron