MIKROBIOLOGIA PRZEMYSŁOWA
SEM VIII
1. Typy odżywiania się mikroorganizmów
2. Podział mikroorganizmów w oparciu o sposób odżywiania się
3. Wyjaśnić pojęcia: fotolitotrofia, chemolitotrofia i organotrofia
4. Oddychanie tlenowe mikroorganizmów
5. Proces fermentacji
6. Charakterystyka bakterii chemolitotroficznych
7. Oddychanie beztlenowe mikroorganizmów
8. Fotosynteza bakteryjna
9. Charakterystyka bakterii prowadzących proces fotosyntezy anoksygenowej
1O.Toksyczny wpływ tlenu na mikroorganizmy
11.Reakcje tlenu podczas przemian mikrobiologicznych
12.Wymagania pokarmowe mikroorganizmów
13.Pienyiastki budulcowe mikroorganizmów
l4 Źródła pierwiastków budulcowych dla mikroorganizmów
15.Podzial mikroorganizmów w oparciu o wymagania pokarmowe
16.Wzrost osobniczy i populacyjny mikroorganizmów
17.Systeniy hodowli drobnoustrojów
18.W drobnoustrojów w hodowli okresowej, fazy wzrostu mikroorganizmów
19.Diauksja
20.Zjawiska shift up” i shift down” w hodowli drobnoustrojów
21.Zmiany w składzie chemicznym drobnoustrojów zależnie od fazy rozwojowej
22.Parametry wzrostu mikroorganizmów w hodowli okresowej
23.Szybkość wzrostu mikroorganizmów, definicja
24.Wzrost ograniczony: limitacja i hamowanie wzrostu mikroorganizmów
25.Wzrost mikroorganizmów w hodowli ciągłej
26zalety i wady hodowli ciągłej
27.Metody mikroskopowe pomiaru ilości drobnoustrojów
28.Metody hodowlane pomiaru ilości drobnoustrojów
29.Tworzenie się biocenoz w środowiskach
30.Sukcesja pierwotna, przykłady
31 .Sukcesja wtórna, przykłady
32.Oddzialywanie mikroorganizmów w środowiskach i bioproduktach
33.Systeniy oddziaływania bezpośredniego
34 systemy oddziaływania pośredniego
35.Oddzialywanie symbiotyczne między mikroorganizmami, przyklady, znaczenie ekologiczne
36.Drapieżnictwo w świecie mikroorganizmów, przykłady, znaczenie biotechnologiczne
37.Systemy oddziaływania pośredniego
38.Komensalizim w świecie mikroorganizmów, przykłady znaczenie biotechnologiczne
39.Protokoopcracyjne powiązania wśród mikroorganizmów, przykłady, znaczenie biotech.
40.Konkurencja i amensalizim jako formy współzależności wśród mikroorganizmów”
1.Typy odżywiania się mikroorganizmów i 2. Podział mikroorganizmów w oparciu o sposób odżywiania się
W procesach wzrostu mikroorganizmy muszą pobierać składniki kom., energię. Energia ta wykorzystywana jest na tzw. Pracę: osmotyczną, chem., fiz. Właściwy sposób odżywiania mikroorganizmów wymaga ustalenia:
a) co jest źródłem energii metabolicznej - energia jest niezbędna komórce dla wykonania pracy chem., np. biosynteza (anabolizm), pracy osmotycznej (transport przez błony), pracy mech. (ruch0, pracy fiz. (świecenie)
b) Co jest źródłem węgla do syntezy składników komórkowych
c) Co jest donorem elektronów i protonów w układzie przenośników, np. transport przez błony i przeminay energetyczne w komórce.
MIKROORGANIZMY dzielimy na: Chemotrofy (chemolitotrofy i chemoorganotrofy) oraz fototrofy
Fototrofy - fotolitotrofy - organizmy samożywne wykorzystujące energie słoneczną. Źródłem węgla jest CO2, źródłem elektronów związki nieorganiczne. Do fototrofów zaliczamy:
A) Beztlenowe bakterie fototropiczne, których proces fotosyntezy nie uwalnia do środowiska tlenu, np. purpurowe bakterie siarkowe
B) Tlenowe mikroorganizmy fototropiczne, takie jak glony czy sinice, które w procesie fotosyntezy wydzielają tlen, takimi właściwościami cechują się sinice i glony.
Chemotrofy - mikroorg., które uzyskują energię w wyniku przemian oksydoredukcyjnych organicznych lub nieorganicznych związków chemicznych. Źródłem węgla dla tej grupy może być CO2 lub zw. organiczne.:
A) Chemolitotrofy - chemoautotrofy - mikroorganizmy korzystające z energii zawartej w zredukowanych zw. nieorganicznych, np. w związkach azotu, siarki, żelaza lub energii zawartej zawartej w H2. Źródłem elektronów mogą być zredukowane związki takie jak: Nh3, CO, H2S, S, Fe3+; źródłem węgla może być CO2 lub zw. nieorganiczne. Do grupy tej zaliczamy bakterie nitryfikacyjne, wodorowe, metanowe, żelaziste i wiele innych, szczególnie często występujących w środowisku wody i gleby
B) Chemoorganotrofy - chemoorganoheterotrofy - organotrofy - mikroorganizmy, które jako źródło węgla, energii i elektronów (donory wodoru) wykorzystują związki organiczne. Należą tutaj wszystkie grzyby, większość bakterii, pierwotniaki oraz zwierzęta.
3. Wyjaśnić pojęcia: fotolitotrofia, chemolitotrofia i organotrofia
Fotolitotrofy - organizmy samożywne wykorzystujące energie słoneczną. Źródłem węgla jest CO2, źródłem elektronów związki nieorganiczne. Do fototrofów zaliczamy:
A) Beztlenowe bakterie fototropiczne, których proces fotosyntezy nie uwalnia do środowiska tlenu, np. purpurowe bakterie siarkowe
B) Tlenowe mikroorganizmy fototropiczne, takie jak glony czy sinice, które w procesie fotosyntezy wydzielają tlen, takimi właściwościami cechują się sinice i glony.
C) Chemoorganotrofy - chemoorganoheterotrofy - organotrofy - mikroorganizmy, które jako źródło węgla, energii i elektronów (donory wodoru) wykorzystują związki organiczne. Należą tutaj wszystkie grzyby, większość bakterii, pierwotniaki oraz zwierzęta.
D) Chemolitotrofy - chemoautotrofy - mikroorganizmy korzystające z energii zawartej w zredukowanych zw. nieorganicznych, np. w związkach azotu, siarki, żelaza lub energii zawartej zawartej w H2. Źródłem elektronów mogą być zredukowane związki takie jak: Nh3, CO, H2S, S, Fe3+; źródłem węgla może być CO2 lub zw. nieorganiczne. Do grupy tej zaliczamy bakterie nitryfikacyjne, wodorowe, metanowe, żelaziste i wiele innych, szczególnie często występujących w środowisku wody i gleby.
Redukcja NAD, (gdy donory wodoru mają wyższy potencjał redox niż NAD) zachodzi na drodze odwróconego transportu elektronów napędzanego przez ATp (NADPH2 jest niezbędny do redukcji CO2) ; elektrony wchodzą do łańcucha oddechowego dopiero na poziomie cyt a i cyt c.
4. Oddychanie tlenowe mikroorganizmów
W oddychaniu tlenowym akceptorem jest tlen, szereg reakcji biochemicznych wymaga obecności tlenu. Drobnoustroje tlenowe mają w swoim wyposażeniu cykl kwasów trójkarboksylowych. Pirogronian jest cyklicznie utleniany w cyklu Krebsa. Pirogronian w wyniku tych przemian ulega całkowitej degradacji do CO2 - to całkowite spalanie zwane jest mineralizacją. W cyklu Krebsa zysk energetyczny jest znikomy, nie kończy on jednak oddychania tlenowego (opisuje los atomów C). Cykl Krebsa przygotowuje dopiero do wytworzenia energii, które ma miejsce w łańcuchu oddechowym. Pełny łańcuch oddechowy funkcjonuje tylko u Eucariota, u Procariota są najczęściej znacznie zredukowane przenośniki elektronów ( np. tylko jeden cytochrom). U Procariota zlokalizowanych jest w błoniach mezosomów a u Eucariota w mitochondriach. Następuje stopniowy odzysk energii. Wśród tlenowców są takie mikroorganizmy, które wykorzystują związki nieorganiczne jako akceptory elektronów (chemolitotrofy). Podczas takiej przemiany dochodzi do utlenienia nieorganicznych związków mineralnych czemu towarzyszy wydzielenie pewnej ilości energii. Grupa tlenowców obejmuje drobnoustroje, dla których obecność tlenu jest niezbędna dla wzrostu. Do tej grupy należy wiele gatunków drożdży i bakterii (Thibacillus, Bacillus, Pseudomonas) oraz większość pleśni. Liczne mikroorganizmy należące do tlenowców są wykorzystywane w procesach biotechnologicznychnp. Do produkcji kwasów organicznych, antybiotyków. Są to pleśnie z rodzaju i Aspergillus, Penicillum, promieniowce. Zapotrzebowanie na tlen hodowli mikroorganizmów jest uzależnione od rodzaju stężenia źródeł węgla i energii, fazy hodowli, gęstości Komorek i ich stanu fizjologicznego.
5. Proces fermentacji
Beztlenowa przemiana pirogronianu do zredukowanego metabolitu, który bez udziału tlenu nie może być dalej metabolizowany, jako produkt niepotzrbny komórce, zostaje wydzielony do środowiska. Znaczącym celem dalszej przemiany kwasu pirogronowego do produktu końcowego jest regeneracja, czyli utlenienie zredukowanego NAD-u podczas II etapu. Proces fermentacji jest charakterystyczny dla org. Względnie beztlenowych - drożdże, bakterie mlekowe oraz bezwzględnych beztlenowców - bakterie z rodzaju Clostridium Bacteroides. Produkty końcowe będą zależne od wyposażenia enzymatycznego. Jeżeli drobnoustroje mają skąpe wyposażenie to powstaje produkt homogeny, np. kwas mlekowy; jeżeli jest ono bogate to układ heterofermentacji.
6. Charakterystyka bakterii chemolitotroficznych
Chemolitotrofy są liczną grupą. Występują w wodzie i glebie głównie. Są to mikroorganizmy, które wykorzystują energię zawarta w zredukowanych związkach chem. I odzyskują te energię w wyniku utleniania związków.
1)Bakterie utleniające wodór - bakterie wodorowe (np. Hydrogenomonas)
H2 + 1/2 O2 H2O + 1ATP
2)Bakterie utleniające amoniak (np. Nitromonas)
NH4+ + 1,5 O2 NO2- + H+ + H2O + 2ATP
3) Bakterie utleniające azotany (np. Nitrobacter)
NH4+ + 1/2O2 NO3- + 1ATP
4)Bakterie utleniające nieorganiczne związki siarki, bakterie siarkowe (np. Thiobacillus)
HS- + H+ ½O2 S0 H2O + 2ATP
S0 + 1½O2 + H2O SO42- +2H+ +1ATP
5) Bakterie utleniające jony żelazowe - bakterie żelaziste ( Ferrobacillus)
Fe2+ +H+ + ¼ O2 Fe3+ + ½ H2O + 1 ATP
7. Oddychanie beztlenowe mikroorganizmów
W przypadku oddychania beztlenowego końcowym akceptorem elektronów mogą być związki organiczne utlenione bądź związki nieorganiczne utlenione, które w wyniku przeniesienia elektronów ulegają redukcji do określonych związków, np. CO2 do CH4. Określony typ oddychania beztlenowego przyjmuje nazwę od związku, który powstaje bądź jest substratem w danym procesie.
Oddychanie beztlenowe ( akceptorem elektronów są utlenione zw. chemiczne) |
|
CO2 CH4 CH3COOOH CH4 |
Węglanowe (bakterie metenogenne) |
S0 H2S |
Siarkowe(Desulforomonas) |
SO42- H2S |
Siarczanowe (Desulfovibrio) |
NO3- NO2-, N2O, N |
Azotanowe (denitryfikatory) |
Fe3+ Fe2+ |
Żelazowe |
8. Fotosynteza bakteryjna
Fotosyntezę bakteryjną można podzielić na akoksygenową (nie powstaje w niej tlen jako produkt fotosyntezy) i oksygenową powstaje tlen). Fotosynteza sinic (cyjanobakterii) jest procesem oksygenowym. W skład tej grupy organizmów zwanych dawniej glonami niebieskozielonymi wchodza formy jednokomórkowe i nitkowate. Aparat fotosyntezy sinic jest zlokalizowany w systemie wewnątrzplazmatycznych błon zwanych tylakoidami, pokrytych fikobilisomami zawierającymi składniki zbierające światło. Barwniki absorbujące światło to chlorofile, które pochłaniają światło czerwone i niebieskie oraz kartenoidy, fikobiliproteiny, fikoerytryna, fikocyjanina, zwane barwnikami dodatkowymi, gdyż absorbują światło o takich długościach fal, przy których chlorofil nie działa wydajnie. Barwniki są uporządkowane w sposób wysoce uporządkowany, co zwiększa efektywność transferu energii świetlnej. Fotosynteza sinic jest podobna do fotosyntezy glonów.
Bakterie zielone i purpurowe tym różnią się między innymi od sinic, że są ścisłymi beztlenowcami i nie wykorzystują wody jako źródła elektronów. Mikroorganizmy te wykorzystuja H2 , H2S i siarkę elementarną jako donory elektronów i mają inne barwniki zwane bakteriochlorofilami, które absorbują światło o większych długościach fali. Wyróżniamy bakteriofil a i b. Bakterie te w czasie fotosyntezy nie wytwarzają tlenu i NADPH + H+. Przeprowadzją cykliczny transport elektronow, który może zostać wykorzystany do syntezy ATP, natomiast nie są zdolne do bezpośredniego wytworzenia NADPH + H+ w czasie fotosyntetycznego przepływu elektrolitów. Zielone bakterie siarkowe mają pewną formę niecyklicznego przepływu elektronów potrzebnego do zredukowania Nad+. Bakterie purpurowe naie dysponuja takim mechanizmem tak, więc NADPH + H+powstaje u nich w wyniku odwrotnego transportu elektronów. W obecności H2 może dojść do bezpośredniego wytwarzania NADPH + H+, jako że H2 ma potencjał redukcyjnybardziej ujemny niż NAD+.
Fotosynteza to przekształcenie energii świetlnej w energię biochemiczną (ATP) oraz siłę redukującą (NADH2).
9. Charakterystyka bakterii prowadzących proces fotosyntezy anoksygenowej
Bakterie fotosyntezy anoksygenowej nie są zdolne do wykorzystywania wody jako donora wodoru, wymagają bardziej zredukowanych donorów, takich jak H2S, H2 lub zw. organiczne. Występują pospolicie w środowisku wodnym, warstwach dennych płytkich stawów, wodach wolno płynących, przy brzegach jezior, w górnych warstwach bagien.
Zawierają bakteriochlorofil a (niektóre również b,d lub e ) oraz karotenoidy (różne zabarwienie komórek).
a) Purpurowe bakterie siarkowe Chromatiaceaejako donory wodoru wykorzystują H2S w komórkach gromadzą ziarna siarki silnie załamujące światło. Różne kształty komórek z reguły dużych rozmiarów ( formy nitkowate 3×20-50 um, kuliste 3-5 um).Są najczęściej ruchliwe
b)Purpurowe bakterie bezsiarkowe Rhodospirillaceae jako donory wodoru wykorzystują związki organiczne H2, rzadko H2S; nie gromadzą siarki w komórkach. Proces fotosyntezy prowadza tylko w warunkach beztlenowych, a w warunkach tlenowych metabolizm heterotroficzny. Komórki małe (kilka um) , pałeczki, formy spiralne, kuliste.
c)Zielone bakterie siarkowe Chlorobiaceae jako donory wodoru wykorzystują H2S, tiosiarczany, H2. Są to nitkowate lub małe pałeczki (0,8-2um), nieruchliwe.
10.Toksyczny wpływ tlenu na mikroorganizmy
Toksyczne działanie tlenu może być:
a)Bezpośrednie, np. utlenianie grupy -SH w białkach, inaktywacje niektórych enzymów, nadoksydacje, cytochromów, nadoksydacje lipidów
b) Pośrednie działanie toksycznych połączeń tlenu.
11.Reakcje tlenu podczas przemian mikrobiologicznych
1)wolne rodniki nadtlenowe(O2):O2+e-→O2
2)nadtlenek wodoru(H2O2):O2+2e-→O2-2
3)rodniki hydroksylowe(OH):O2+H2O+H+→O2+H2O+OH
Reakcja 1 jest katalizowana przez oksydazę cytochromową i niektóre enzymy zawierające miedz. Reakcja 2 jest charakt. Dla niektórych enzymów z gr.flawinową (oksydazę: glukozowa, aminokw, ksantynowa).Reakcja 3 jest katalizowana przez oksydazy:ksantynowa,aldehydowa.
Ochronne działanie wykazują enzymy:
2H2O2→2H2O+O2 katalaza
H2O2+NADH→2H2O+NAD-peroksydaza
2O2+2H+→H2O2+O2dysmutaza nadtlenkowa
4O2-+4H+→2H2O+3O2dysmutaza nadtlenowa(katalaza)
Prowadząc hodowle mikrorg tlenowych należy pamiętać, że, pomimo iż tlen jest niezbędnym związkiem dla ich wydajnego metabolizmu, zbyt wysokie stężenie może być jednak niebezpieczne(toksyczne połączenie tlenu).
12.Wymagania pokarmowe mikroorganizmów
Wszystkie mikroorganizmy wymagają do wzrostu tych samych pierwiastków, ale w różnych formach. Pierwiastki budulcowe niezbędne to: C, N,O,H,P,S. Wchodzą one w skład podst. związków chemicznych budulcowych i funkcjonalnych. Opracowując skład pożywek hodowlanych musimy pamiętać o tym, że liczbowo reakcja tych składników musi zabezpieczać podst wymagania na składniki budulcowe. Najprostszą metodą na zbadanie tych wymagań jest zbadanie biomasy po hodowli. C,H,O podst. składniki związków org., budujących składniki funkcjonalne, np. enzymy. Komórka najchętniej wykorzystuje źródła węgla, które nie wymagają wstępnej hydrolizy i bez transformacji bezpośrednio wprowadzone są do szlaków metabolicznych. Mikroorg potrafią wykorzystać większość źródeł węgla, ale muszą wtedy zaangażować część energii w procesy hydrolizy i transformacji wprowadzonych związków.
13.Głównne składniki masy komórkowej mikroorganizmów
Do gł. składników masy komórkowej należą:
węgiel do 50%,wodór do 8%, tlen do 30%, azot do 14%, fosfor do 3%, siarka do 1%
Te 6 związków jest podst. związkami org., które nazywa się również pierwiastkami biogennymi.
14. Źródła pierwiastków budulcowych mikroorganizmy
Do źródeł pierwiastków budulcowych zaliczamy: O, C, H, N, P ,S oraz składniki funkcjonale.
TLEN- jego źródłem dla mikroorg beztlenowych są związki organiczne, dla tlenowców i względnych tlenowców powietrze.
WODÓR-wykorzystywany jest wodór pochodzący z połączeń organicznych i wody, która jest wszechobecna w środowisku hodowlanym.
AZOT-jest składnikiem aminokw., zasad azotowych. Źródłem azotu dla mikroorg. Są jony NH4+,najchętniej wykorzystywane. Część mikroorg potrafi wykorzystywać azot w formie utlenionej, ale wbudowany może być tyko w formie zredukowanej. Dlatego tracona jest energia podczas redukcji. Istnieją mikroorg. zdolne do wykorzystywania mineralnych źródeł azotu: większość bakterii chorobotwórczych, bakterie fermentacji mlekowej. Muszą mieć one dostarczany azot w postaci aminokwasów lub białek. Nieliczne drobnoustroje zdolne są do wykorzystania azotu atmosferycznego (Azotobacter).
FOSFOR-wchodzi w skład fosfolipidów (składniki błon komórkowych), kwasów nukleinowych, ATP, AMP,ADP. Źródłem fosforu są związki mineralne, najczęściej PO3-4. Mikroorg potrafią też wykorzystywać fosfor w połączeniach organicznych.
SIARKA-wyst. w organizmie w formie zredukowanej. Mikroorg. najchętniej pobierają siarkę w postaci utlenionej i tylko niektóre drobnoustr. nie potrafią tej siarki redukować, wtedy dostarcza się im S2-.
SKŁ. FUNKCJONALNE-muszą być również obecne w pożywce. Biorą udział w przemianach metabolicznych. Najistotniejsze są dla mikroorg.:Na,Mg,Ca,Fe. Związki te wykorzystywane są z reguły w postaci rozpuszczalnych soli. Wymagane są również pierwiastki śladowe gł. Mn, Mo, Zn, Cu, Co, Ni, Se. Pierwiastki te w pożywkach w skład, których wchodzą składniki naturalne, tzw. pożywki przemysłowe są pomijane gdyż wyst w zanieczyszczeniach surowców, w wodzie.
15.Podział mikroorg w oparciu o wymagania pokarmowe.
Ze względu na wymagania pokarmowe mikroorg dzielimy na:
a)O skrajnie wysokich wymaganiach pokarmowych(mikroogr. chorobotwórcze).Rozwijają się w środowisku o pełnym składzie pokarmowym(pełny zestaw budulcowy związków organicznych).Najlepiej rozwijają się w organizmach żywych, czyli w środowisku identycznym jak ich właściwa materia komórkowa. Na skraju tych wymagań są niektóre gatunki bakterii mlekowych.
b)O skrajnie niskich wymaganiach pokarmowych(autotrofy węglowe, azotowe, niektóre sinice).Rosną w środowiskach bardzo ubogich, są charakterystyczne dla wód i gleby.
c)O pośrednich wymaganiach pokarmowych, występują w przyrodzie najliczniej.
16.Wzrost osobniczy i populacyjny mikroorg.
a)Jako przyrost biomasy i objętości organizmu jednokomórkowca np. komórki drożdżowej lub bakteryjnej lub wielokomórkowca np. strzępki pleśniowej w wyniku biocenozy substancji komórkowych tj.: białka. kw. nukleinowe, nukleotydy, służących do budowy struktur komórkowych: jądra, mitochondria, rybosomów, błon elementarnych i innych.
b) Jako wzrost populacji tzn. zwiększenie biomasy populacji drobnoustrojów rozumianej jako zbiór organizmów określonego gatunku (szczepu) znajdujących się w danym środowisku np.. w hodowli.
W biotechnologii mikrobiologicznej i mikrobiologii żywności wzrost drobnoustrojów opisany jest przede wszystkim jako wzrost populacji.
Obserwacje wzrostu populacji bakterii i drożdży pozwalają na dostrzeżenie istotnych różnic w charakterze tego procesu dla obu wymienionych grup jednokomórkowców. Komórka macierzysta dzieli się na 2 komórki potomne. Zatem w danym momencie hodowli bakteryjnej obecne są tylko rówieśnicze komórki tej samej generacji. Odmiennie jest w hodowli drożdży, gdzie komórka macierzysta po wypączkowaniu z niej komórki potomnej zdolna jest do dalszego rozmnażania. Konsekwencją takiego sposobu rozmnażania jest jednoczesna obecność w hodowli komórek z różnych pokoleń, przy czym komórki pochodzące z różnych generacji mogą być równocześnie, zaś komórki siostrzane mogą się różnić od siebie wiekiem.
Czas generacji jest to czas niezbędny do powstawania nowego pokolenia komórek. Znając czas trwania hodowli t i liczbę pokoleń wygenerowanych w tym czasie n można obliczyć czas generacji z następnej zależności. g=t/n gdzie t- czas trwania hodowli, liczba pokoleń wegetatywnych w tym czasie
rys.modelowy wzrost populacji bakterii
g
Komórka macierzysta→komórka macierzysta+ komórka potomna
↑
substraty odżywcze
*→linia generacyjna najstarszej komórki potomnej
y,z→rówieśnicze komórki z różnych generacji
x→najstarsza komórka potomna
x,z→siostrzane komórki potomne w różnym wieku
rys.modelowy wzrost drożdży
Systemy hodowli mikroorganizmów
System otwarty
Składniki pokarmowe są w sposób stały wprowadzane do naczynia hodowlanego, zatem w naczyniu panuje stałe stężenie składników hodowli przy jednoczesnym odprowadzaniu przefermentowanego podłoża wraz z produktami metabolizmu, które są dla mikroorg niekorzystne. Jest to hodowla ciągła.
System zamknięty(hodowla okresowa)
Jest to system hodowli wznawianej. Mikroorg namnażają się, osiągają pewną gęstość, hodowla zatrzymuje się, aby hodowla dalej trwała musimy przenieś mikroorg na świeżą pożywkę. Mikroorg poprzez ciągłą zmianę składników odżywczych pożywki napotyka na zmienne warunki hodowli mikroorg następuje zmiana chemicznych składników w medium hodowlanym. Produkty końcowe są tu nagromadzone jednocześnie z ubytkiem składników odżywczych pożywki. Ilość tych produktów zależy od uzdolnień enzymatycznych komórki, czyli szlaków składających się na metabolizm komórki dostarczającej energię. Produkty są często toksyczne, mikroorganizmów, ponieważ nie ma ich odprowadzania następuje wzrost stężenia tych produktów-to prowadzi do samozatrucia hodowli, która zaczyna zamierać. Hodowla okresowa to taka hodowla, mikroorganizmów której komórki w każdym momencie trwania hodowli mają odmienny skład chemiczny środowiska na skutek ubytku substancji odżywczych, nagromadzenia metabolitów toksycznych.
18.Wzrost mikroorganizmów w hodowli okresowej fazy wzrostu mikroorganizmów
Krzywa wzrostu drobnoustrojów w hodowli okresowej: 1-faza adaptacja, 2-faza wzrostu logarytmicznego, 3-faza zwolnionego wzrostu, 4-faza stacjonarna, 5-faza zwolnionego
Faza 1, nazywana fazą zastoju, faza adaptacyjna lag fazą, trwa od momentu wprowadzenia drobnoustroju do świeżej pożywki, aż do czasu uzyskania intensywnego rozwoju. Okres jej trwania zależy od składu i rodzaju pożywki hodowlanej drobnoustrojów, temperatury inkubacji. W fazie zastoju masa pojedynczych komórek wprowadzonych do świeżej pożywki rośnie natychmiast w stosunku logarytmicznymi.
Faza 2- faza wzrostu logarytmicznego (wykładnicza) charakteryzuje się najintensywniejszym przyrostem liczby komórek, który odbywa się ze stałą szybkością. Masa i liczba komórek wzrastają logarytmicznie. Natomiast wartości RNA w przeliczeniu na masę komórek oraz masa komórki są względnie stałe. Składniki komórki są syntetyzowane w sposób uporządkowany i ze stałą szybkością co sprzyja zachowaniu stałej wielkości komórki oraz jej składu chemicznego. Czas trwania fazy wykładniczej jest determinowany czynnikami środowiskowymi, zależy też od właściwości drobnoustrojów i sposobu prowadzenia hodowli. Faza 3 jest fazą zwolnionego wzrostu, podczas której maleje rozmiar komórek oraz stają się one ubogie w RNA i rybosomy. Wzrasta liczba komórek martwych.
Faza 4 czyli faza stacjonarna, cechuje się stałą liczbą komórek w hodowli jako wynik równowagi pomiędzy liczbą komórek żywych i martwych. Ze względu na ograniczoną ilość dostępnego substratu komórki zużywają materiały zapasowe, ulegają również degradacji części rybosomów. Komórki jednak nadal zachowują zdolność do syntezy różnych enzymów. Jest to faza produkcji idiolitów.
Faza 5 jest to faza zwolnionego zamierania, następuje przyrost komórek martwych, pojawiają się formy inowulacujne oraz zachodzą procesy autolizy. Faza 6 jest to faza logarytmicznego zamierania. Szybkość zamierania i okres trwania tej fazy zależy od stopnia wrażliwości mikroorganizmów na toksyczne metabolity występujące wówczas w dużych stężeniach.
19. Diauksja
Funkcjonowanie represji katabolicznej indukcji substratowej sprawia, że w obecności dwóch źródeł energii, drobnoustroje w pierwszej kolejności wykorzystują substrat metabolicznie korzystniejszy ==, a dopiero po jego wyczerpani następuje indukcja dodatkowych enzymów i uruchomienie nowego odcinka procesu katabolicznego umożliwiającego komórce przyswajanie substratu gorszego. Właśnie ten mechanizm stanowi podstawę zjawiska , „Diauksja” czyli dwufazowość wzrostu drobnoustrojów. Ma on również kluczowe znaczenie w wielu procesach biosyntezy. Przykładem może być biosynteza antybiotyków, która wymaga ograniczenia szybkości wzrostu drobnoustrojów. Można to uzyskać obniżając stężenie łatwo przyswajalnego cukru Np. glukozy, do poziomu niezbędnego do szybkiego namnożenia odpowiedniej ilości komórek i wprowadzając do podłoża dodatkowy, wolniej przyswajalny cukier, Np. laktozę, sprzyjający efektywnej produkcji antybiotyku, klasycznym przykładem takiego procesu jest biosynteza penicyliny.
20. Zmiany w składzie chemicznym drobnoustrojów zależności od fazy rozwojowej
1 - zawartość RNA w komórce,2- Sucha masa komórek w 1cm3 hodowli, 3- liczba komórek w 1 cm3, 4- masa pojedynczej komórki a- początek fazy wzrostu logarytmicznego, b- czas wyczerpania się substratu, c- koniec wykładniczego wzrostu komórek, d- początek fazy stacjonarnej.
W fazie 1 zawartość RNA zwiększa się nawet 12- krotnie, wzrasta ilość białka oraz rybosomów. Ostatecznie powiększają się wymiary pojedynczych komórek. Masa poszczególnych komórek oraz zawartość RNA w komórce rośnie do osiągnięcia pewnego poziomu. W fazie logarytmicznej masa i liczba komórek wzrastają logarytmicznie. Natomiast wartość RNA w przeliczeniu na masę komórki są względnie stałe. Składniki komórki są syntetyzowane w sposób uporządkowany i ze stała szybkością, co sprzyja zachowaniu stałej wielkości komórki oraz jej składu chemicznego. Faza III jest fazą zwolnionego wzrostu podczas której maleje rozmiar komórek oraz stają się one ubogie w RNA i rybosomy .Wzrasta liczba komórek martwych .Ze względu na ograniczoną ilośc dostępnego substratu komórki zużywają materiały zapasowe ulegają również degradacji części rybosomów.
21. Zmiany w składzie chemicznym drobnoustrojów zależności od fazy rozwojowej
1-zawartość RNA w komórce,2- Sucha masa komórek w 1cm3 hodowli, 3- liczba komórek w 1 cm3, 4- masa pojedynczej komórki a- początek fazy wzrostu logarytmicznego, b- czas wyczerpania się substratu, c- koniec wykładniczego wzrostu komórek, d- początek fazy stacjonarnej.
W fazie 1 zawartość RNA zwiększa się nawet 12- krotnie, wzrasta ilość białka oraz rybosomów. Ostatecznie powiększają się wymiary pojedynczych komórek. Masa poszczególnych komórek oraz zawartość RNA w komórce rośnie do osiągnięcia pewnego poziomu. W fazie logarytmicznej masa i liczba komórek wzrastają logarytmicznie. Natomiast wartość RNA w przeliczeniu na masę komórki są względnie stałe. Składniki komórki są syntetyzowane w sposób uporządkowany i ze stała szybkością, co sprzyja zachowaniu stałej wielkości komórki oraz jej składu chemicznego. Faza III jest fazą zwolnionego wzrostu podczas której maleje rozmiar komórek oraz stają się one ubogie w RNA i rybosomy .Wzrasta liczba komórek martwych .Ze względu na ograniczoną ilośc dostępnego substratu komórki zużywają materiały zapasowe ulegają również degradacji części rybosomów.
22. Parametry wzrostu mikroorganizmów w hodowli okresowej
Wzrost drobnoustrojów w hodowli okresowej mogą scharakteryzować trzy parametry krzywej wzrostu zdefiniowane przez Monado, tj przyrost biomasy, szybkość wzrostu i czas trwania fazy zastoju
przyrost Biomasy (rys1)
plan
przyrost biomasy
szybkość wzrostu
czas trwania fazy zastoju
wyd produktu
czas trwania fazy zastoju (rys2)
Parametry wzrostu w hodowli okresowej: A - przyrost biomasy, B- czas trwania fazy zastoju
Przyrost biomasy stanowi różnicę pomiędzy ilością biomasy w szczytowym punkcie hodowli a ilością biomasy wprowadzonej do pożywki czyli : X=Xmax-X0. Zależność ta określa wartość suchej masy w gramach. Największy przyrost biomasy przypada na fazę logarytmicznego wzrostu, lecz największą ilość biomasy stwierdza się w fazie stacjonarnej. Jest ona plonem rozwoju populacji we wszarskich fazach wzrostu hodowli okresowej.
Szczególne znaczenia dla charakterystyki hodowli okresowej ma zależność wyrażona stosunkiem przyrostu bi0omasy i zużycia substratu, zwana współczynnikiem wydajności lub wydajnością wzrostu , oznaczona symbolem Y, (g s.m. komórek/g substratu). Wydajność może być również odniesiona do molowego stężenia substratu, wówczas wyraża molowy współczynnik wydajności Ym. Molarny współczynnik wydajności umożliwia obliczenie ilości ATP otrzymanej z danego substratu.
23. Szybkość wzrostu mikroorganizmów- definicja
Szybkość właściwa wzrostu μ, jest podstawowym parametrem charakteryzującym wzrost drobnoustrojów w różnych warunkach ( zarówno organicznych jak i nieorganicznych), Jest ona definiowana jako stosunek przyrostu liczby komórek istniejących, N lub jako stosunek szybkości przyrostu masy komórkowej do biomasy już istniejącej X:
![]()
lub ![]()
μ- szybkość właściwa wzrostu
N - (liczba kom. Już istniejących) N - gęstość kom (liczba kom. W jednostce objętości)
dN - przyrost liczby kom.
dx - przyrost masy kom
X- biomasa już istniejąca
Parametrem równoważnym μ jest okres generacji, tg, definiowany jako czas potrzebny do podwojenia liczby komórek. Zależność pomiędzy tymi dwoma parametrami jest następująca:
![]()
tg - okres generacji - czas potrzebny do 2x kom
Jeżeli wzrost populacji wyrażany jest przez zmianę stężenia biomasy, to równanie szybkości wzrostu nieorganicznego ma postać:
![]()
równanie szybkości wzrostu nieorganicznego
![]()
- max szybkość wzrostu
X - biomasa już istn.
24. Wzrost organiczny - limitacja i hamowanie wzrostu mikroorganizmów.
Często w warunkach laboratoryjnych i przemysłowych mamy do czynienia ze zjawiskiem limitacji wzrostu przez niskie stężenie któregoś ze składników podłoża lub hamowania wzrostu w obecności inhibitora, np. gromadzonego metabolitu. W hodowli okresowej zaistnienie takich warunków organicznych szybkość wzrostu prowadzi di zakończenia fazy laboratoryjnej, mikroorganizmów zatem wzrost organiczny przestaje być w tym wypadku wzrostem wykładniczym
Obniżenie się stężenia jednego ze składników podłoża powoduje zwykle postępujący spadek szybkości wzrostu zgodnie z modelem Monoda analogicznym do równania Michaelita- Menten:
![]()
Gdzie: S- stężenie substratu, Ks- stała półnasycenia, czyli stężenie substratu przy którym μ=1/2 μmax
Na podstawie powyższego równania łatwo zauważyć, ze![]()
Jeżeli substrat asymilowany przez drobnoustroje znajduje się w zbyt dużym nadmiarze, może to również prowadzić do ograniczenia szybkości wzrostu do takich substratów należą: kwas octowy, etanol - stosowane w niektórych biotechnologiach.
Rys zależność między szybkością właściwą wzrostu drobnoustrojów, μ a stężeniem S składnika pożywki:
1 - składnik limitujący, 2 - składnik limitujący ( w niskim stężeniu) i hamujący wzrost ( w wysokim stężeniu)
Ważnym czynnikiem ograniczającym wzrost drobnoustrojów jest niedobór tlenu w środowisku. W warunkach niedostatecznego natlenienia szybkość wzrostu i szybkość oddychania biomasy zależą od stężenia tlenu rozpuszczonego w podłożu.
Częstym czynnikiem hamującym szybkość wzrostu jest nagromadzenie toksycznego produktu metabolizmu: 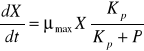
P- stężenie produktu
25. Wzrost mikroorganizmów w hodowli ciągłej.
W hodowli ciągłej drobnoustrojów po osiągnięciu przez mikroorganizmy wybranej fazy wzrostu następuje stałe zasilanie hodowli strumieniem świeżej pożywki. Jednocześnie z fermentora jest odprowadzana ta sama ilość podłoża zawierającego podłoże i szkodliwe metabolity . Zatem objętość hodowli w czasie trwania procesu pozostaje niezmienna. Z tego wynika, że do utrzymania stałego stężenia biomasy w hodowli ciągłej niezbędna jest równowaga dynamiczna pomiędzy biomasą wypłukiwaną, a namnażaną. Procesy ciągłe wykorzystywane są najczęściej do otrzymywania białka paszowego, do produkcji etanolu, butanolu, acetonu, kwasów organicznych, niektórych antybiotyków oraz przy oczyszczaniu ścieków metodą osadu czynnego.
Rozróżnia się dwa sposoby prowadzenia hodowli ciągłej:
na zasadzie chemostatu
W chemistacie wzrost drobnoustrojów jest regulowany szybkością dopływu pożywki. W dowolnym udziale można zmienić szybkość przepływu pożywki tylko w takim zakresie aby stężenie wprowadzonego substraty nie przekroczyło wartości, przy której drobnoustroje osiągają maksymalną właściwą szybkość wzrostu. W chemostacie stałe stężenie biomasy jest wynikiem stałej szybkości rozcieńczania D
na zasadzie turbidostatu
W metodzie tej zadane stężenie biomasy utrzymywane jest w wyniku automatycznej regulacji przepływu podłoża opartej na pomiarze zmętnienia zawiesiny hodowlanej. W skonstruowanym układzie między natężeniem przepływu pożywki a zmętnieniem zawiesiny hodowlanej działa mechanizm ujemnego sprzężenia zwartego tzn. gdy zmętnienie maleje natężenie przepływu wzrasta i odwrotnie - wzrostowi zmętnienia towarzyszy zmniejszenie dopływu świeżej pożywki. Wadą hodowli w turbidostacie jest niższy niż w chemostacie stopień wykorzystywania substratu oraz konieczność stosowania skomplikowanych technicznie urządzeń
Rys.
Hodowla ciągła:
a - zapas jałowej pożywki
h - naczynie hodowlane
p - pompa dozująca pożywkę
z - odbieralnik nadmiaru hodowli
o - rurka syfonowa odprowadzająca nadmiar hodowli
26. Zalety i wady hodowli ciągłej
Zalety:
wyeliminowanie wpływu czasu na warunki i fizjologię drobnoustrojów
możliwość prowadzenia hodowli w dowolnie długim czasie w warunkach ustalonych
możliwość regulacji stanu fizjologicznego drobnoustrojów przez dobór szybkości zasilania substratem i dobór składu pożywki zasilającej hodowlę
jednorodność stanu fizycznego i chemicznego hodowli
możliwość automatyzacji procesu
większa szybkość i wydajność wielu procesów
efektywniejsze wykorzystanie aparatury
Wady:
niebezpieczeństwo degradacji szczepów i opanowania hodowli przez mutanty o niekorzystnych własnościach technologicznych
trudności z utrzymaniem aseptycznych warunków w bioreaktorze
niekorzystne wskaźniki reologiczne wynikające z tworzenia przez niektóre drobnoustroje wielokomórkowych agregatów, kłaczków, zarastanie ścian fermentora i przewodów
niesprzyjające warunki produkcji niektórych substancji wytwarzanych przez komórki nierosnące
niedostateczna znajomość dynamicznych właściwości drobnoustrojów w hodowli ciągłej
27. metody mikroskopowe pomiaru ilości drobnoustrojów
Metody mikroskopowe polegają na bezpośrednim liczeniu komórek mikroorganizmów pod mikroskopem. Różnice sprowadzają się tylko do sposobu przygotowania próby i różnych metod przeliczania drobnoustrojów na jednostkę objętości:
liczenie drobnoustrojów przy użyciu komór
komory do liczenia drobnoustrojów pod mikroskopem maja postać szklanych płytek z wyciętym wgłębieniem podzielonym na kwadraty lub prostokąty o znanej powierzchni
KOMORA THOMA
Ma ona głębokość 0,1mm na jej dnie znajduje się siatka składjąca się z 16 dużych kwadratów, które z kolei są podzielone na 16 małych. Szerokość o długość komory wynosi 0,05mm. Na powierzchni komory thoma wyżłobione są trzy kanaliki w kształcie litery H. Badana zawiesinę nanosi się na górną i dolną siateczkę w centralnej części komory. Wyjściowa gęstość zawiesiny powinna wynosić od 106 do 107 komorek w 1cm3 podczas liczenia wyznacza się średnia liczbę komórek w około 40 małych kwadracikach. Liczbę drobnoustrojów w 1cm3 wylicza się ze wzoru:
L = 4*106*n*a
n - rozcieńczenie zawiesiny drobnoustrojów
a - średnia zawartość komórek w polu widzenia.
KOMORA BURKERA
Ma podobną budowę jak komora Thoma, podzielona jest jednak na większe kwadraty: o boku 0,2mm. Liczbę drobnoustrojów wylicza się ze wzoru:
L = 2,5*105*n*a
n - rozcieńczenie zawiesiny drobnoustrojów
a - średnia zawartość komórek w polu widzenia.
Liczenie drobnoustrojów w preparacje przeżyciowym (bezpośrednim)
Polega ona na wykonaniu preparatu bezpośredniego a następnie liczeniu pod mikroskopem średniej liczby komórek. Liczenie należy przeprowadzić w trzech preparatach minimum po 20 pól widzenia w każdym. Należy tez policzyć powierzchnię pola widzenia mikroskopu. Liczbę drobnoustroju w 1cm3 próby oblicza się z wzoru:
![]()
n - rozcieńczenie zawiesiny drobnoustrojów
a - średnia zawartość komórek w polu widzenia
r - promień pola widzenia [mm]
h - grubość warstwy cieczy między szkiełkami [mm]
Liczba drobnoustrojów metodą Breeda w preparacie barwionym
Zasada polega na wykonaniu równomiernego rozmazu niewielkiej ilości hodowli, pobranej tzw. pipetą Breeda (0,01cm3), na znanej powierzchni A szkiełka podstawowego, wcześniej dokładnie odtłuszczonego. Po wykonaniu rozmazu, wysuszeniu i utrwaleniu preparat wybarwia się odpowiednim barwnikiem a następnie liczy pod mikroskopem średnią liczbę komórek z trzech preparatów w 20 wybranych polach widzenia. Mikrometrem obiektywowym wyznacza się średnicę pola wodzenia. Liczbę drobnoustrojów w 1cm3 próby oblicza się z wzoru:
![]()
pole rozmazy
a - średnia zawartośc komórek w polu widzenia
x- = 100, przelicznik dla pipety Breeda
r - promień pola widzenia
Metoda ta może byś stosowana do liczenia bakterii i grzybów
Metoda DELT
Polega na liczeniu pod mikroskopem drobnoustrojów na filtrze membranowym o porach 0,45μm , po uprzednim ich wybarwieniu fluorochromami. Badane próbki mogą być poddane wstępnej obróbce enzymatycznej. Po filtracji komórki barwi się oranżem akrydyny i liczy w mikroskopie fluorescencyjnym. Barwienie, umożliwia odróżnianie komórek żywych od martwych; żywe komórki fluoryzują na pomarańczowo lub żółto, martwe na zielono.
28. Metody hodowlane pomiaru ilości drobnoustrojów
Metody hodowlane pośrednie oparte SA na zdolności drobnoustrojów do rozmnażania, dzięki czemu oznacza się tylko żywe komórki zdolne do wzrostu w pożywkach płynnych ( metoda rozcieńczeniowa) lub w pożywkach stałych ( metoda płytkowa i jej modyfikacje)
Metoda rozcieńczeniowa
Metoda seryjnych rozcieńczeń Listera należy do klasyczny technik stosowanych do określania liczby drobnoustrojów oraz izolowania czystych hodowli ze środowiska płynnego. Zasada tej metody polega na wieloetapowym rozcieńczaniu badanej zawiesiny tak aby w 1cm3 znajdowała się 1 komórka. Z kolejnych rozcieńczeń wykonuje się posiewy, po1cm3, do pożywek płynnych, co najmniej w 2 powtórzeniach. Po inkubacji określa się ilość prób dodatnich tj. takich w którym rosną drobnoustroje. Korzystając z tablicy McCrady'ego ujmujących statycznie ilość porób ze wzrostem drobnoustrojów, rozcieńczeń i ilością powtórzeń wylicza się w oparciu i rachunek prawdopodobieństwa NPL, czyli najbardziej prawdopodobna liczbę drobnoustrojów w 1 cm3 badanej proby. Dokładność metody zależy od przygotowania właściwych rozcieńczeń i od ilości równoległych prób. Warunkiem jest przygotowanie takiego szeregu rozcieńczeń, aby w ostatnim nie były już obecne mikroorganizmy. Jest to metoda czasochłonna, pracochłonna i coraz zadziej stosowana.
Metoda płytkowa
Zasada metody polega na wysiewie odpowiednio rozcieńczonej zawiesiny drobnoustrojów na pożywkę stałą, inkubacji i liczeniu wyrosłych koloni. Wyniki otrzymywane tą metodą są zawsze niższe niż rzeczywiste, ponieważ liczy się na płytkach tylko kolonie tych drobnoustrojów, które SA zdolne do wzrostu na danej pożywce i w danych warunkach. Dodatkowym czynnikiem obniżającym wyniki oznaczeń jest występowanie niektórych bakterii w skupiskach i łańcuszkach. Technika wysiewu na płytki wymaga zachowania następujących procedur:
ROZCIEŃCZENIE ZAWIESINY
Badaną próbę należy tak rozcieńczyć, aby po wysiewie na płytkę wyrosło od 30-300 kolonii. Rozcieńczenia najczęściej przygotowuje się jałowej soli fizjologicznej lub płynie Ringera.
POSIEW NA PŁYTKACH PETRIEGO
Posiew można wykonywać metoda wgłębna lub powierzchniowa. Powinny być wykonywane z dużych rozieńczeń, w co najmniej dwóch powtórzeniach
INKUBACJA
Płytki z posiewem inkubuje się w temperaturze optymalnej l wzrostu określonej grupy mikroorganizmów
LICZENIE
Po inkubacji liczy się wyrosłe kolonie na płytkach, odrzucając płytki z ilością powyżej 300
Modyfikacja metody płytkowej
Tradycyjna metoda liczenia drobnoustrojów na pożywce stałej doczekała się wielu modyfikacji, które w znacznym stopniu ja uproszczają. Automatyzacja podstawowych etapów, takich jak: rozcieńczanie badanej próby, posiew i liczenie drobnoustrojów zmniejsza praco - i czasochłonność wykonywanych badań.
Metoda filtrów membranowych
Jest stosowana do określania ilości drobnoustrojów w środowiskach, przeważnie wodnych, w których ilość ich jest niewielka, poniżej 20-30 komórek w 1cm3. Zasada tej metody polega na przesączeniu określonej objętości badanego płynu (lub rozpuszczonej substancji stałej) przez filtr membranowy o wielkości porów 0,2-0,4μm. Filtracja odbywa się dzięki wytworzeniu podciśnienia w kolbie ssawkowej za pomocą pompki wodnej lub mechanicznej. Ilość cieczy przeznaczonej do filtracji dobiera się według przewidywanej liczby mikroorganizmów w badanej próbie. Zatrzymane na filtrze drobnoustroje rozwijają się poprzez umieszczenie filtru na powierzchni płytki Petriego z odpowiednią pożywką agarową. Płytki inkubuje się w temperaturze optymalnej dla wzrostu danych mikroorganizmów w czasie 24*-48godzin. Po inkubacji ilość kolonii, które wyrosły na powierzchni filtru odpowiada liczbie drobnoustrojów znajdujących się w badanej objętości płynu.
Wykorzystanie Petrefilmów
Petrifilmy są jałowymi plastykowymi płytkami zawierającymi gotowe pożywki hodowlane przykryte folią polietylenową. Specjalny indykator dodany do pożywek zabarwia kolonie drobnoustrojów przez co są one lepiej widoczne. Na płytkach są zaznaczone kwadraty o powierzchni 1cm3 ułatwiające liczenie wyrosłych kolonii. Posiewu dokonuje się za pomocą pipety nanosząc 1cm3 badanej próbki lub jej rozcieńczenia na środek płytki, następnie przykrywa się folią i dociska specjalnym krążkiem w celu równomiernego rozprowadzenia próbki po całej powierzchni pożywki. Po inkubacji płytek w określonych warunkach liczy się wyrosłe kolonie.
Płytki kontaktowe testy łopatkowe
Testy te SA przeznaczone do szybkiej oceny jakości mikrobiologicznej produktów oraz kontroli stanu higienicznego. Zastosowano w nich technikę wzrostu drobnoustrojów na pożywce agarowej nałożonej na obydwie strony płytki, która jest umieszczona w sterylnej fiolce. Producenci oferują zestawy do określenia ogólnej liczby drobnoustrojów, liczby drożdży i pleśni, obecności bakterii z rodzaju Enterobacteriaceae, identyfikacji bakterii E. coli, oraz bakterii z rodzaju Pseudomonas.
Metoda posiewów spiralnych
W metodzie tej następuje rozprowadzenie mikropipetą badanego materiału po powierzchni płytki agarowej w postaci spirali Archimedesa, biegnącej spiralnie od centrum płytki do jej brzegów. W urządzeniu służącym do posiewów tą metodą silnik elektryczny obraca pustawe, w której umieszcza się otwartą płytkę Petriego z pożywką agarową. Liczenie drobnoustrojów odbywa się przez umieszczenie pod płytką wzorcowej siatki podzielonej na 8 sektorów, będących wycinkami koła.
29. Tworzenie się biocenoz w środowiskach..
Niezależnie od poglądu ze mikroorganizmy znajdziemy wszędzie, są środowiska ich pozbawione. Są to np. tereny objęte spływem lawy, świeżo odkryte powierzchnie skał, tkanki żywych , zdrowych organizmów wyższych czy surowce spożywcze i produkty wyjałowione. Takie środowiska są doskonałymi modelowymi warunkami do obserwowania tworzenia się biocenzo. Stwierdzono, ze w podobnych warunkach ekologicznych kolejność zmian wykazuje pewną prawidłowość. Zjawiska tez nazwano sukcesją ekologiczną. Wyróżniamy dwa typy sukcesji ekologicznej: pierwotną i wtórną (patrz pyt 30 o 31)
30. Sukcesja pierwotna - przykłady
W ekosystemach w miarę upływu czasu zmienia się stan gatunkowy roślin i zwierząt oraz warunki środowiskowe. Zmiany w ekosystemie mogą być antropogeniczne ( wywołane działalnością człowieka) oraz naturalne. Zmiany naturalne mogą być sezonowe, odwracalne. Mogą wystąpić zmiany naturalne wieloletnie, nazywane sukcesjami. Sukcesje są procesami nieodwracalnymi, trwającymi do ustalenia pełnej równowagi między asymilacją (produkcja), a dysymilacją (konsumpcją). Równowaga osiągana jest w stadium nazywanym klimaksem (najbardziej stabilne studium sukcesji). Sukcesja jest procesem kierunkowych zmian biocenozy, powodujących przeobrażenie się prostych ekosystemów w bardziej złożone. Mechanizm sukcesji polega na tym, że organizmu przekształcając środowisko, w czasie bytowania w nim, czynią je przydatnym do innych organizmów.
Sukcesja pierwotna - występuje na terenach dziewiczych, pozbawionych jakichkolwiek organizmów. Miejsca objęte sukcesją pierwotną to: wydma, skała, hałda, zatopiony statek. Sukcesja pierwotna jest właściwa dla środowisk, w których dotąd nie istniało Zycie, np. na świeżo odkrytych skalach. Rozwój biocenzo w takich środowiskach zaczyna się od gatunków pierwotnych, czyli tzw. mikroorganizmów pionierskich, które stopniowo zmieniają środowisko , przez co staje się bogatsze w różne gatunki i w ich odrębnie w ilości osobników.
Gatunki pionierskie zazwyczaj cechują się zdolnością ruchu lub zdolnością do przeżycia w powietrzu. Daje im to szansę dotarcia do określonego siedliska. Rodzaj gatunków pionierskich, zależy od charakteru chemicznego i parametrów fizycznych zasiedlanego środowiska. W środowiskach jałowych o bardzo ubogim składzie odżywczym w warunkach sukcesji pierwotnej pierwszymi gatunkami mikroorganizmów są producenci, tj. mikroorganizmy fotoautotroficzne i chemolitotroficzne o bardzo małych wymaganiach pokarmowych. Najczęściej mikroorganizmami pionierskimi w takich środowiskach są sinice lun glony. Biomasa tych organizmów po śmierci tanowi doskonały, pełnowartościowy materiał budulcowy, dzięki któremu mogą pojawić się reducenci i konsumenci, czyli mikroorganizmy chemoorganotroficzne. Ze względu na z reguły duża szybkość wzrostu bakterie chemoorganotroficzne stają się okresowo dominującymi w populacji. Jednakże po wyczerpaniu nagromadzonych składników pokarmowych może dojść do ponownego powrotu organizmów autotroficznych.
W środowiskach jałowych, lecz bogatych odżywczo, np. po wprowadzeniu zanieczyszczeń organicznych do wód czy też w tkankach zwierząt po uboju, czy otwarciu jałowych produktów lub surowców spożywczych jako pierwsze pojawiają się mikroorganizmy organotroficzne, wyspecjalizowane w zdolności wykorzystywanie określonych odżywczych składników chemicznych. Jeżeli jest to środowisko bogate, lecz homogenie składem, wtedy zespół mikroorganizmów pionierskich jest bogaty ilościowo, ale jednorodny gatunkowo. W przypadku środowiska o zróżnicowanym składzie chemicznym obserwuje się biocenozy o bogatym układzie gatunkowym. Jako pierwsze rozwijają się gatunki o najmniejszej liczebności i najwyższej szybkości wzrostu. Kolejno rozwijają się gatunki wolniej rosnące lub wykorzystujące produkty metabolizmu poprzedników.
31. Sukcesja wtórna - przykłady
W ekosystemach w miarę upływu czasu zmienia się stan gatunkowy roślin i zwierząt oraz warunki środowiskowe. Zmiany w ekosystemie mogą być antropogeniczne (wywołane działalnością człowieka) oraz naturalne. Zmiany naturalne mogą być sezonowe, odwracalne. Mogą wystąpić zmiany naturalne wieloletnie, nazwane sukcesjami. Sukcesje są procesami nieodwracalnymi, trwającymi do ustalenia pełnej równowagi między asymilacja (produkcją) o dysymilacja (konsumpcją) Równowaga osiągana jest w stadium nazywanym klimaksem (najbardziej stabilne studium sukcesji). Sukcesja jest procesem kierunkowych zmian biocenozy, powodujących przeobrażenie się prostych ekosystemów w bardziej złożone. Mechanizm sukcesji polega na tym, że organizmu przekształcając środowisko, w czasie bytowania w nim, czynią je przydatnym do innych organizmów.
Sukcesja wtórna - występuje na miejscu zniszczonego ekosystemu lub na obszarach zajętych przez inna biocenozę. Sukcesja wtórna zachodzi o wiele szybciej niż pierwotna (od kilku do kilkudziesięciu lat). Miejscem występowania sukcesji wtórnej jest: pozbawione upraw pole, zarastający staw, pozostawiony bez opieki trawnik.
Sukcesja wtórna polega na rozwinięciu biocenoz mikroorganizmów z pominięciem fazy pionierskiej. Takie tworzenie się biocenoz występuje w środowiskach, w których zakłócono istniejący zespół na skutek klęski żywiołowej lub ingerencji człowieka. Przykładem może być zbiornik wody, w którym została zniszczona naturalna biocenoza na skutek suszy czy suszenia wody. W przypadku działalności człowieka w przetwórstwie spożywczym może to być pasteryzacja surowca lub produktu. W wyniku tego zabiegu dochodzi do zabicia części mikroorganizmów, czyli usunięcia części ekosystemu. Takie środowiska są z reguły bogate w składniki odżywcze, co pozwala na równomierny rozwój różnych grup mikroorganizmów, mikroorganizmów układzie jednak z reguły niespecyficznym dla składu chemicznego danego środowiska
32. Oddziaływanie mikroorganizmów w środowiskach i bioproduktach.
O układzie drobnoustrojów poza czynnikami środowiskowymi decydują również wzajemne stosunki pomiędzy poszczególnymi gatunkami biocenozy. Wzajemne oddziaływanie między mikroorganizmami podzielić można na:
Bezpośrednie (symbioza , pasożytnictwo, drapieżnictwo)
Pośrednie (protokooperacja, komensalizm, konkurencja, amensalizm)
Populacje występujące w biocenozie mogą na siebie wzajemnie oddziaływać. Interakcje mogą być protekcyjne lub antagonistyczne. Przy braku zdolności mówimy o neutralizmie. Do zależności protekcyjnych, nazywanych nieantagonistycznymi, zaliczamy:
Symbiozę - która jest rodzajem współżycia organizmów czerpiących obopólne korzyści, np. bakterie brodawkowe i korzenie roślin motylkowych. Jeśli występuje rodzaj współżycia tzw. koniecznego, wówczas ten układ nazywamy mutualizmem - przykładem są porosty, których ciało zbudowane jest z glonów i strzępek grzyba.
Komensalizm - jeden z występujących organizmów w układzie jest kornensalem, czerpiącym korzyść z obecności drugiego osobnika tzw. gospodarza, który nie ponosi szkód, np. porosty występujące na pniach drzew.
Protokooperacja - dotyczy dwóch organizmów świadczących sobie wzajemne usługi, „korzyści”, ale nie jest to konieczne do ich egzystencji.
Do zależności antagonistycznych zaliczamy:
Konkurencję - występuje wówczas gdy są w danym siedlisku populacje o podobnych wymaganiach życiowych ( np. podobne sposoby odżywiania, jednakowe wymagania środowiskowe). W konkurencji wygrywa populacja liczebniejsza lub mająca większe umiejętności przystosowawcze.
Pasożytnictwo - polega na wykorzystaniu organizmu żywiciela przez pasożyta. Wyróżniamy pasożyty zewnętrzne i wewnętrzne. Pasożyty wytworzyły wiele cech przystosowujących do pasożytnictwa, np. narządy czepne, oskórki chroniące przed strawieniem , doskonałe rozmnażanie.
Drapieżnictwo - dotyczy sytuacji, w której osobnik jednego gatunku (drapieżnik) chwyta, zabija i zjada osobniki drugiego gatunku (ofiara) Drapieżca w stosunku do ofiary jest zwykle większy (gdy jest mniejszy poluje stadnie). Zabija się zwykle osobniki młode, stare, słabe, chore. Ilość osobników drapieżców jest ściśle uzależniona od ilości ofiar. Drapieżcy posiadają szereg przystosowań ułatwiających zdobycie pożywienia (dobry węch, wzrok, rozwinięte kły, pazury, ewentualnie dzioby) a ofiary do obrony przed pożarciem (barwa ochronna, szybkie nogi, czujność)
Amensalizm - występuje wówczas, gdy czynności jednej populacji szkodzą innym, np. tworzone przez bobry żeremia zmieniają warunki wodne w biocenozie, wykluczając obecność dotychczasowych populacji.
Antybioza - wytwarzanie antybiotyków ( związków chemicznych) przez jedną grupę bakterii powoduje zahamowanie wzrostu innej.
33. System oddziaływania bezpośredniego.
Symbioza
Symbiozą nazywamy taki rodzaj współzależności dwóch lub więcej różnych gatunków ściśle od siebie zależnych które bez obecności partnera rozwijają się bardzo słabo lub wcale nie rosną, Taki rodzaj współzależności jest również nazywany mutualizmem lub symbiozą mutualiztyczną. Znane są przykłady oddziaływania symbiotycznego miedzy samymi mikroorganizmami, jak i między mikroorganizmami i organizmami wyższymi w tym również człowiekiem. Głownie kierunki korzystnego oddziaływania na siebie symbiontów są wynikiem
Wymiany składników pokarmowych
Przekształcania przez mikrosymbioty nieoprzysfajalnych dla organizmu wyższego substancji pokarmowych:
Dostarczanie substancji wzrostowych
Zaopatrywanie w składniki mineralne
Wykorzystywanie i w ten sposób usuwania, produktów metabolizmu toksycznych dla organizmu partnera
Ochrony przed szkodliwymi czynnikami środowiskowymi
Zmiany parametrów środowiska
Pasożytnictwo
Jest rozumiane jako współzależność, w której jeden z partnerów (pasożyt) osiąga korzyści, natomiast drugi partner(gospodarz)nie ponosi lub ponosi szkody. W pierwszym przypadku pasożytnictwo dotyczy rozkładu martwych szczątków roślin czy zwierząt. Ten rodzaj pasożytnictwa jest szeroko rozpowszechniony w przyrodzie i jest decydujący dla zapewniania obiegu pierwiastków. Drugi rodzaj pasożytnictwa występuje wtedy, gdy gospodarzem jest organizm żywy. W świecie mikroorganizmów tego rodzaju pasożytnictwo jest stosunkowo mało poznane. Przykładem mogą być bakteriofagi atakujące komórki bakterii.
Drapieżnictwo
Jest systemem, który rozumiany jest najczęściej jako odżywianie się jednych mikroorganizmów innymi. W świecie zwierząt jest to system współzależności bardzo często spotykany, natomiast między mikroorganizmami należy do rzadkości. Najbardziej typowym przykładem pasożytnictwa u mikroorganizmów jest odżywianie się pierwotniaków bakteriami. Jest to zjawisko szczególnie widocznie w zbiornikach wodnych, osadach czynnych , ściekach. Główną rolą w eliminowaniu bakterii ściekowych przypisuje się orzęskom i wiciowcom.
34 System oddziaływania pośredniego.
W przyrodzie współzależności między i zachodzące poprzez środowisko występują niezwykle często Warunkiem niezbędnym jest jednak odpowiednio bliskie sąsiedztwo organizmów, tak, aby tworzone metabolity czy zmiany parametrów fizycznych środowiska mogły wywierać wpływ na partnerów.
Protokooperacja
Jest często określana jako pośrednia symbioza. Jest to system, w którym wszystkie powiązane ze sobą mikroorganizmy odnoszą korzyści W tym systemie nie ma konieczności współistnienia jednakże wspólne bytowanie jest korzystne dla partnerów i objawia się zwiększeniem szybkości wzrostu i, wyższą aktywnością metaboliczną większą ekspansywnością w środowisku lub większą tolerancją na zmienione warunki bytowania.
Współzależności protokooperacyjne polegają na:
-Wzajemnym uprzystępnianiu składników pokarmowych
- Wzajemnej wymianie gazów, najczęściej dotyczy to, CO i 02
- Wytwarzaniu i wzajemnej wymianie substancji wzrostowych przez partnerów zespołu.
- Wytwarzanie substancji stymulujących wzrost i usuwanie metabolitów toksycznych przez współbytując mikroorganizmy.
Komensalizm
Oznacza współzależność, w wyniku której jeden z partnerów odnosi korzyści, natomiast drugi nie podlega wpływowi, istnienie partnera jest dla niego obojętne Jest to tzw jednostronna korzyść z reguły tego typu zależności są w małym stopniu swoiste Komensalizm najczęściej polega na:
-Przeprowadzeniu przez jednego z mikroorganizmów substratów pokarmowych,
nieprzyswajalnych przez partnera w produkty, które juz może wykorzystać jako
składniki odżywcze;
-Tworzeniu przez jednego ze współmieszkańców ekosystemu substancji wzorcowych,
np. witamin, stymulujących wzrost partnerów;
-Rozkładzie lub wykorzystywaniu w środowisku substancji hamujących wzrost
partnerów;
Konkurencja
Jest formą współrzycia w której obydwaj partnerzy współzawadniczą o deficytowy i ważny dla nich składnik pokarmowy bądź też o światło, wodę czy przestrzeń życiową Konkurencja występuje tylko w takich przypadkach, gdy zasoby substancji potrzebnej dla rozwoju obydwu grup są zbyt małe, aby zabezpieczyć potrzeby współistniejących mikroorganizmów. współzawodniczące mikroorganizmy nie szkodzą sobie nawzajem lecz walczą o zaspokojenie własnych potrzeb.
Amensalizm
Często określany jest antagonizmem; jest formą współzależności w wyniku, której rozwój jednej populacji jest zahamowany przez substancje wytwarzane przez partnera. Wytwarzanie w tym środowisku drapieżnictwo pierwotniaków jest uznawane jako efekt korzystny, pozwalający na redukcje substancji antagonistycznych może być korzystne dla wytwarzającego je mikroorganizmu Osłabianie szybkości wzrostu wrażliwych partnerów lub ich eliminowanie daje producentowi szansę uzyskania przewagi w ekosystemie i ekspansji środowiska Jest to szczególnie istotne dla mikroorganizmów wolno rosnących, które mają małe możliwości konkurowania z intymni mieszkańcami biocenozy. Często substancje
antagonistyczne są traktowane jako „broń” mikroorganizmów w walce o przetrwanie w środowisku.
35.Oddziaływanie symbiotyczne między mikroorganizmami, przykłady, znaczenie ekologiczne.
Symbiozą nazywamy taki rodzaj współzależności dwóch lub więcej różnych gatunków ściśle od siebie zależnych, które bez obecności partnera rozwijają się bardzo słabo lub wcale me rosną Taki rodzaj współzależności jest również nazywany mutualizmem lub symbiozą mutualiztyczną. Znane są przykłady oddziaływania symbiotycznego miedzy samymi mikroorganizmami, jak i miedzy mikroorganizmami i organizmami wyższymi w tym również człowiekiem Główne kierunki korzystnego oddziaływania na siebie symbiontów są wynikiem:
-Wymiany składników pokarmowych,
- Przekształcania przez mikrosymbionty nieprzyswajalnych dla organizmu wyższego substancji pokarmowych;
- Dostarczania substancji wzrostowych,
- Zaopatrywania w składniki mineralne,
-Wykorzystywania i w ten sposób usuwania, produktów metabolizmu toksycznych dla organizmu partnera;
-Ochrony przed szkodliwymi czynnikami środowiskowymi,
-Zmiany parametrów środowiska
Typowym przykładem oddziaływania symbiotycznego wśród mikroorganizmów są zespoły porostów złożone z układów glonów lub sinic z grzybami Zewnętrzna struktura porostów w małym stopniu przypomina budową partnerów W poroście glon i grzyb są tak ściśle powiązane ze sobą ze stanowią jeden organizm wegetatywny, przy czym grzybnia oplata całkowicie komórki glonów W i niektórych przypadkach strzępki grzybni mogą wnikać do wnętrza komórek glonów.
Uważa się, ze zapoczątkowanie symbiozy zachodzi zazwyczaj przy niesprzyjających warunkach środowiska np. na suchym podłożu zbyt intensywnym świetle. Z reguły porosty kolonizują ekstremalne ekosystemy, w których żaden z partnerów nie może istnieć samodzielnie. Glony zaopatrują, komórki grzybów w organiczne substancje pokarmowe natomiast grzyby dostarczają im soli mineralnych oraz chronią przed niekorzystnymi warunkami środowiska. Niektóre pierwotniaki mogą zawierać wewnątrz komórek fotoautotroficzne glony spełniające rolę symbiontów. Szczególnie wyraźnie występuje taka zależność pomiędzy rzęskami a. zielenicami. Ciekawym przykładem symbiozy jest współzależność pomiędzy glonami i bakteriami, np. pałeczki z rodzaju Pseudomonas mogą rozwijać się w cytoplazmie Volvox. Pałeczki mogą również występować w komórkach pierwotniaków Uważa się, ze odgrywają rolę w trawieniu pewnych składników pokarmowych nieprzyswajalnych przez pierwotniaki Symbiotyczne zależności mogą łączyć również partnera mikrobiologicznego z organizmem wyższym. Szczególne znaczenie ma symbioza pomiędzy roślinami motylkowymi i bakteriami z rodzaju Rhizobium Bardzo rozwinięty jest system symbiotycznej zależności pomiędzy mikroorganizmami a zwierzętarni, w tym człowiekiem Takim ekosystemem gdzie ta zależność występuje szczególnie wyraźnie jest przewód pokarmowy Zwierzęta żywiące się wyłącznie materiałem roślinnym bogatym w błonnik zawierają w przewodzie pokarmowym mikroorganizmy degradujące ten sacharyd Najciekawszy zespół mikroorganizmów występuje u przeżuwaczy Żwacz ich w zasadzie jest jedną wielka komorą fermentacyjną Mikroorganizmy żwacza są bardzo specyficzne i poza tym środowiskiem praktycznie nie występują w środowisku.
Mikroflora jelitowa stanowi również bardzo złożony ekosystem, w skład. którego wchodzi ok 500 różnych gatunków mikroorganizmów. korzystna funkcja obecnych w jelicie mikroorganizmów polega na syntezie witamin głównie z grupy B współuczestniczeniu w trawieniu składników pokarmowych oraz na ochronie człowieka przed nadmiernym rozwojem topogenów jelitowych.
36. Drapieżnictwo w świecie mikroorganizmów, przykłady, znaczenie biotechnologiczne.
Jest system który rozumiany jest najczęściej jako odżywianie się jednych mikroorganizmów innymi. W świecie zwierząt jest to system współzależności bardzo często
spotykany, natomiast między mikroorganizmami należy do rzadkości Najbardziej typowym przykładem pasożytnictwa u mikroorganizmów jest odżywianie się pierwotniaków bakteriami Jest to zjawisko szczególnie widoczne w zbiornikach wodnych, osadach czynnych, ściekach W tym środowisku drapieżnictwo pierwotniaków jest uznawane jako efekt korzystny, pozwalający na redukcję ilości osadu czynnego Główną rolą w eliminowaniu bakterii ściekowych przypisuje się orzęskom i wiciowcom Obecność i odpowiednia ilość pierwotniaków w osadzie czynnym jest uznawana jako wskaźnik dobrze skojarzonej biocenozy Podobne zależności można również spotkać w glebie, gdzie pierwotniaki będą żywiły się bakteriami do najpowszechniej spotykanych drapieżców będą tu należały wiciowce i ameby Zależność drapieżca — ofiara występuje również między pierwotniakami i bakteriami w żołądku z przeżuwających.
38. Komensalizm w świecie mikroorganizmów, przykłady, znaczenie biotechnologiczne.
Oznacza współzależność, w wyniku której jeden z partnerów odnosi korzyści natomiast drugi nie podlega wpływowi, istnienie partnera jest dla niego obojętne Jest to tzw. jednostronna korzyść z reguły tego typu zależności są w małym stopniu swoiste. Komensalizm najczęściej polega na:
-Przeprowadzeniu przez jednego z mikroorganizmów substratów pokarmowych nieprzyswajalnych przez partnera, w produkty, które nie może wykorzystać jako składniki
W wodzie i glebie ten rodzaj zależności jest dość powszechny i najczęściej polega na rozkładzie sacharydów lub białek do produktów łatwo przyswajalnych przez partnerów;
Mikroorganizmy glebowe, niezdolne do wykorzystywania takich sacharydów jak celuloza czy hemicelulozy, zależne są od grzybów wydzielających do środowiska enzymy hydrolityczne rozkładające te substraty. Podobnie niektóre mikroorganizmy zdolne do wykorzystywania aminokwasów jako źródła azotu, zależne są od obecności bakterii proteolitycznych czyniących białka przyswajalnymi dla partnerów.
-Tworzenie przez jednego ze współmieszkańców ekosystemu substancji wzrostowych np. witamin stymulujących wzrost partnerów,
-Rozkładzie lub wykorzystywaniu w środowisku, substancji hamujących wzrost partnerów.
Przykładem może być zależność komensalna między mikroorganizmami tlenowymi i beztlenowymi polegająca na wykorzystaniu tlenu przez mikroorganizmy tlenowe i w ten sposób umożliwienie wzrostu beztlenowcom Zależność ta została wykorzystana w hodowli bakterii beztlenowych metodą Fortnera Korzystne warunki mogą być stworzone również przez zmniejszenie lub podwyższenie pH środowiska Przykładem jest wykorzystywanie kwasów organicznych, prze co stwarzane są korzystne warunki dla wzrostu partnerów wrażliwych na obecność tych kwasów Silnie toksyczny H2S wytwarzany przez bakterie
proteolityczne podczas rozkładu białek, wykorzystywany jest przez bakterie siarkowe które utleniają go do wolnej siarki.
W środowiskach naturalnych lub spożywczych o bogatym składzie chemicznym, często zależności komensalne mają charakter wielostopniowy Wówczas zwane jest to metabiozą lub sukcesją.
39 protokooperacyjne powiązania w świecie mikroorganizmów, przykłady znaczenie biotechnologiczne
Jest często określana jako pośrednia symbioza Jest to system w którym wszystkie powiązane ze sobą mikroorganizmy odnoszą korzyści. W tym systemie nie ma konieczności współistnienia, jednakże wspólne bytowanie jest korzystne dla partnerów i objawia się zwiększeniem szybkości wzrostu, wyższą aktywnością metaboliczną większą ekspansywnością w środowisku lub większą tolerancją na zmienione warniki bytowania
Współzależności protokooperacyjne polegają na
-Wzajemnym udostępnianiu składników pokarmowych
Przykładem może być zespół złożony z bakterii celulolitycznych i bakterii asymilujących azot atmosferyczny, np. z rodzaju Azotobacter Bakterie te dostarczają partnerom zredukowanych, przyswajalnych związków azot, natomiast bakterie celulolityczne, degradując celulozę, zaopatrują zespół w łatwo przyswajalne źródło węgla (glukozę)
-wzajemnej wymianie gazów najczęściej dotyczy to CO2 i O2
Taka zależność łączy heterotroficzne bakterie tlenowe i glony w ściekach Podczas mineralizacji związków organicznych bakterie wydzielają duże ilości CO2 który jest wykorzystywany przez fotoautotroficzne glony i sinice jako źródło węgla, glony natomiast
wyniku metabolizmu fotosyntetycznego zaopatrują partnerów w tlen Ten rodzaj współzależności występuje powszechne w wodzie i podczas oczyszczania ścieków W podobnym systemie zależne są od siebie bakterie beztlenowe i tlenowe w glebie
-wytwarzaniu i wzajemnej wymianie substancji wzrostowych przez partnerów zespołu.
Tego rodzaju zależności występują między bakteriami jogurtowymi. W początkowym okresie hodowli środowisko opanowują szybciej rosnące paciorkowce produkując poza kwasem mlekowym również kwas octowy, aldehyd octowy, kwas mrówkowy, którego obecność jak również obniżony potencjał oksydoredukcyjny środowiska sprzyjają rozwojowi pałeczek jogurtowych. Pałeczki jogurtowe cechują się wyższą niż paciorkowce aktywnością proteolityczną i uwalniają niskocząsteczkowe peptydy i aminokwasy z białek co stymuluje rozwój słabo proteolitycznych szczepów streptococcus thermophilus.
-Wytwarzanie substancji stymulujących wzrost i usuwanie metabolitów toksycznych przez współbytujące organizmy
przykładem może być zespół bakterii fermentujących cukry z wytworzeniem substancji organicznych (substancja toksyczna) z grzybami wykorzystującymi kwasy organiczne jako źródło węgla co jest korzystne dla obydwu partnerów współbytujących w środowisku. Ten system współzależności łączy bakterie fermentacji mlekowej z grzybami Geotrichum candidum czy Candida mycoderma i jest powszechny w produktach fermentowanych, szczególnie roślinnych czy w mleku.
40 Konkurencja i amensalizm jako forma współżycia wśród mikroorganizmów.
Konkurencja
jeśli formą współżycia, w której obydwaj partnerzy współzawodniczą o deficytowy i ważny dla nich składnik pokarmowy bądź też o światło, wodę czy przestrzeń życiową Konkurencja występuje tylko w takich przypadkach, gdy zasoby substancji potrzebnej dla rozwoju obydwu
grup są zbyt małe, aby zabezpieczyć potrzeby mikroorganizmów Współzawodniczące mikroorganizmy nie szkodzą sobie nawzajem, lecz walczą o zaspokojenie własnych potrzeb. Szansa wygrania walki konkurencyjnej jest głównie uzależniona od:
- Szybkości wzrostu i namnażania
-wydajności na czynniki środowiska
-wydajności energetycznej podczas metabolizowania składników pokarmowych;
-wymagań w stosunku do substancji wzrostowych;
- Zdolności do gromadzenia substancji zapasowych i wykorzystywania ich, gdy środowisko ubożeje;
-Zdolności do ruchu lub rozrastania się w postaci strzępek, czyli zdolności do tzw. ekspansji środowiska,
Najostrzejsza konkurencja występuje między mikroorganizmami o podobnych parametrach wzrostu i podobnych wymaganiach pokarmowych Sinice będą konkurowały o światło i CO2 z glonami, promieniowce o tlen i składniki odżywcze z pleśniami
Konkurencja o przestrzeń życiową jest stosunkowo rzadka i dotyczy głównie zależności między mikroorganizmami tlenowymi porastającymi powierzchnię
• Amensalizm
Często określany jest antagonizmem jest formą współzależności, w wyniku, której rozwój jednej populacji jest zahamowany przez substancje wytwarzane przez partnera W tym środowisku drapieżnictwo pierwotniaków jest uznawane jako efekt korzystny, pozwalający na redukcje substancji antagonistycznych może być korzystne dla wytwarzającego je mikroorganizmu Osłabianie szybkości wzrostu wrażliwych partnerów lub ich eliminowanie daje producentowi szanse uzyskana przewagi w ekosystemie i ekspansji środowiska Jest to szczególnie istotne dla mikroorganizmów wolno rosnących, które mają małe możliwości konkurowania z innymi mieszkańcami biocenozy Często substancje antagonistyczne są traktowane jako „bron” mikroorganizmów w walce o przetrwanie środowisku. Zjawisko amensaliznu jest bardzo powszechnie wykorzystywane do utrwalana surowców i produktów spożywczych Tworzone na drodze mikrobiologicznej kwasy organiczne, a szczególnie mlekowy, octowy, propionowy zwiększają stabilność biologiczną żywności fermentowanej Obniżenie pH na skutek rozwoju bakterii fermentacji mlekowej hamuje rozwój wielu bakterii w tym również chorobotwórczych Kwas propionowy jest silnym inhibitorem rozwoju grzybów. Kwas octowy produkowany z etanolu przez bakterie octowe zabija większość mikroorganizmów w środowisku ich występowania jak również nie pozwala na osiedlanie się nowych. W biokonserwacji żywności czynnikiem antagonistycznym działającym na mikroorganizmy zanieczyszczające jest również H2O2 wiele mikroorganizmów fermentujących w tym również bakterie fermentacji mlekowej z rodzaju Lactobacillus są zdolne do wytwarzania H2O2 szczególnie wrażliwe na ten metabolit są bakterie beztlenowe, z rodzaju Clostridium Wiele mikroorganizmów jest znanych ze zdolności do produkcji antybiotyków .Są one silnymi substancjami antydrobnoustrojowymi (antybioza) W środowisku występowania bakterii siarkowych z rodzaju Thiobacillus obserwuje się całkowite zahamowanie wzrostu innych mikroorganizmów.
15
Wyszukiwarka
Podobne podstrony:
ściąga opracowane pytania z pierwszego koła, Biotechnologia środowiska, PŁ, BiNoŻ, studia mgr II sto
Wyniki kolokwium z części spirytusowo-drożdżowej, Biotechnologia środowiska, PŁ, BiNoŻ, studia mgr I
Enzymologia poprawiona sciaga, Biotechnologia środowiska, PŁ, BiNoŻ, studia mgr II stopnia, Semestr
2010 2011biokonwersja kol, Biotechnologia środowiska, PŁ, BiNoŻ, studia mgr II stopnia, Semestr II,
Pytania ogĂllne ochrona środowiska studia stacjonarne II stopnia
Czasy miedzyzielone III-I, Studia Mgr, II semestr mgr, System sterowania ruchem
Tomanek 11.05.2011, Studia Mgr, II semestr mgr, Zarządzanie Przedsięwzięciem Budowlanym
Gleboznawstwo leśne studia mgr drugiego stopnia
PNN materiały, Pielęgniarstwo UM łódź, studia mgr, II semestr, piel. nefrologiczne
Czasy miedzyzielone II-III faza, Studia Mgr, II semestr mgr, System sterowania ruchem
wyklad 11 Istota i spoleczne znaczenie organizacji, studia mgr rok 1, I rok semestr I, współczesne d
Wyklad 2, Studia Mgr, II semestr mgr, Mechanistyczne metody wymiarowania nawierzchni
TN materiały, Pielęgniarstwo UM łódź, studia mgr, II semestr, piel. nefrologiczne
HD materiały, Pielęgniarstwo UM łódź, studia mgr, II semestr, piel. nefrologiczne
Czas pracy kierowców w PL i UE, STUDIA - Kierunek Transport, STOPIEŃ I, SEMESTR 6, Organizacja przew
Rozporządzenie Rady numer 1083, studia mgr rok 1, I rok semestr I, Fndusze strukturalne
Wyklad 5-6, Studia Mgr, II semestr mgr, Mechanistyczne metody wymiarowania nawierzchni
Wyklad 3-4, Studia Mgr, II semestr mgr, Mechanistyczne metody wymiarowania nawierzchni
więcej podobnych podstron