
REGULACJA EKSPRESJI GENÓW EUKARIOTYCZNYCH
1) Etapy regulacji ekspresji genów - przegl
ą
d
2) Regulacja bezpo
ś
rednia przez czynniki zewn
ę
trzne
3) Regulacja po
ś
rednia poprzez czynniki zewn
ę
trzne
4) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– typ kojarzeniowy dro
ż
d
ż
y
5) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów –
powstawanie immunoglobulin
6) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów –
powstawanie makronukleusa orz
ę
sków

1) Etapy regulacji ekspresji genów - transkrypcja
A) Efekt chromatyny - zale
ż
no
ść
intensywno
ś
ci trankrypcji od
regionalnej struktury chromatyny
- Wyciszanie rejonów chromosomów znakowanych przez
modyfikacje zasad (np. przez metylacj
ę
)
- Wyciszanie rejonów poprzez lncRNA (nast
ę
pny slajd)
- Wyciszanie rejonów poprzez lncRNA (nast
ę
pny slajd)
- Pozycjonowanie nukleosomów (kolejny slajd)
B) Wła
ś
ciwa transkrypcja
- Efekt odsłoni
ę
cia DNA przez nadmiarow
ą
(?) transkrypcj
ę
- Specyficzna inicjacja transkrypcji zale
ż
na od sekwencji
promotora i czynników białkowych (aktywatory i represory)
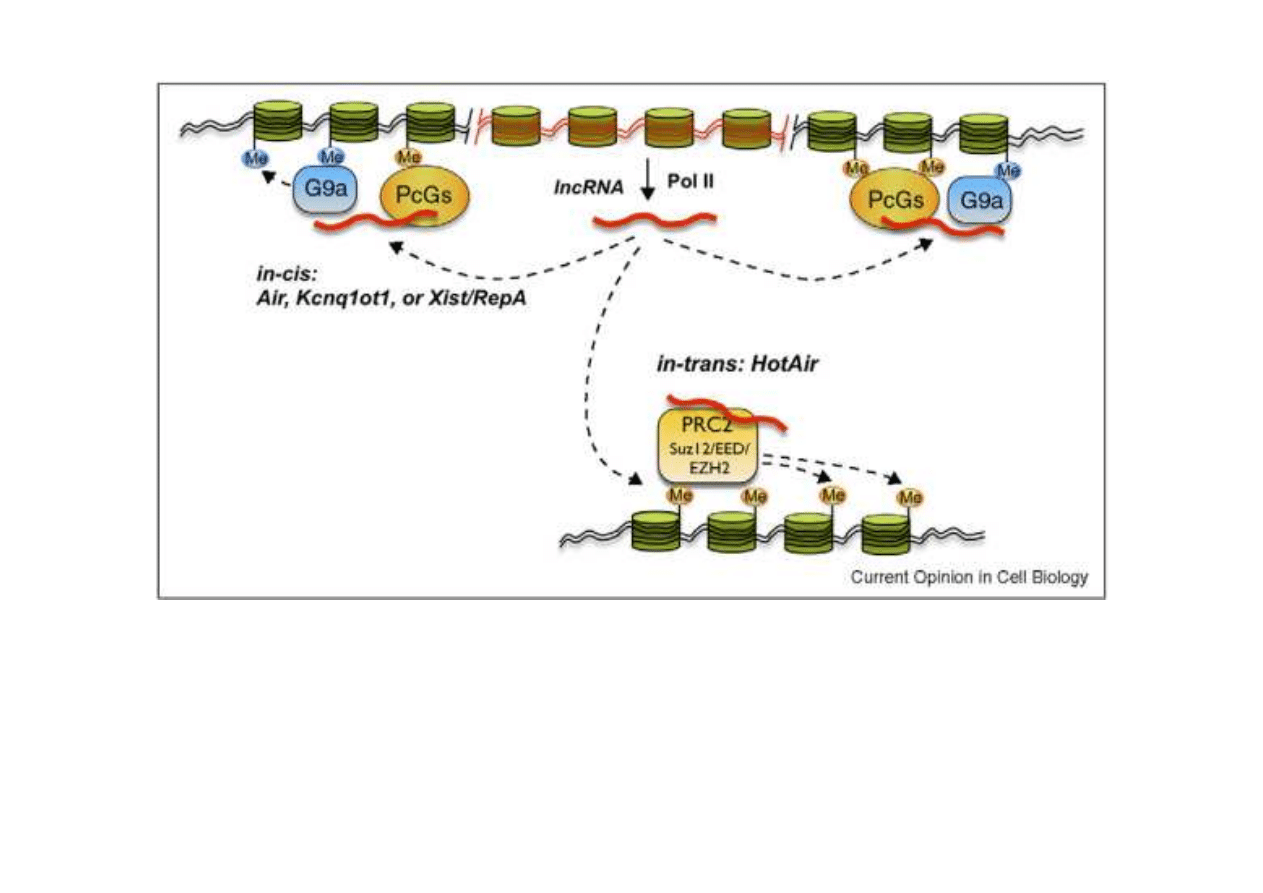
Przykład modyfikacji chromatyny przez RNA. Długi niekoduj
ą
cy RNA (long
non-coding, lncRNA) jest syntetyzowany przez polimeraz
ę
RNA II. Ł
ą
czy
si
ę
z białkami np. G9a i PcGs, które mog
ą
zag
ę
szcza
ć
chromatyn
ę
w
bliskiej okolicy (in cis) lub z innymi białkami, np. PRC2, które prowadz
ą
do
zag
ę
szczenia chromatyny w rejonach odległych (in trans). U ssaków
poznano ju
ż
kilka tysi
ę
cy ró
ż
nych lncRNA.
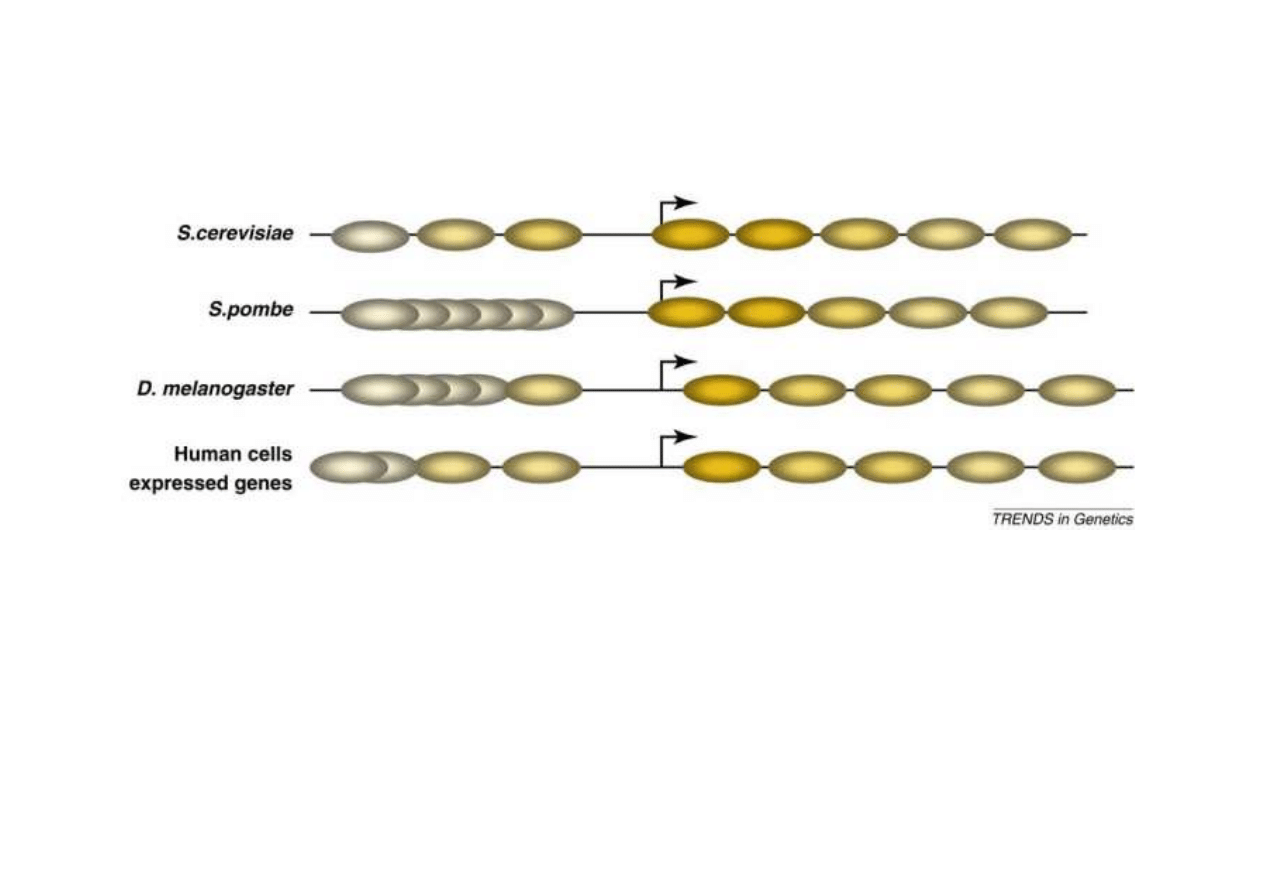
Nukleosomy s
ą
g
ę
sto uło
ż
one (pozycjonowne) w rejonach nie koduj
ą
cych a
najrzadsze w rejonach promotorowych (by ułatwi
ć
inicjacje transkrypcji). W
rejonach koduj
ą
cych s
ą
uło
ż
one w specjalny regularny sposób, tak by ich
uwalnianie i wi
ą
zanie było sprawne.

1) Etapy regulacji ekspresji genów – obróbka mRNA
- Dodawanie czapeczki – sposób regulacji spotykany w
dojrzewaniu jaj u zwierz
ą
t
- Poliadenylacja – alternatywne miejsca dodawania ła
ń
cuchów
poliadenylowych reguluj
ą
np. kwitnienie u ro
ś
lin
poliadenylowych reguluj
ą
np. kwitnienie u ro
ś
lin
- Alternatywne składanie egzonów - bardzo powszechne
- Degradacja dojrzałego mRNA - np. gdy małe odcinki RNA,
tzw. miRNA, odszukuj
ą
komplementarne mRNA co przywołuje
białka degraduj
ą
ce RNA(interferencja RNA)

1) Etapy regulacji ekspresji genów – synteza i obróbka białek
- Inicjacja translacji – np. fosforylacja czynnika translacji elF-2
spowalnia tworzenie kompleksów inicjacyjnych
- Alternatywne ci
ę
cie polipeptydów – pozwala na modyfikacje
białek w zale
ż
no
ś
ci od miejsca syntezy (tkanki)
białek w zale
ż
no
ś
ci od miejsca syntezy (tkanki)
- Modyfikowanie chemiczne – np. u białek sygnałowych cz
ę
sta jest
fosforylacja, u strukturalnych acetylacja
- Selektywna degradacja białek
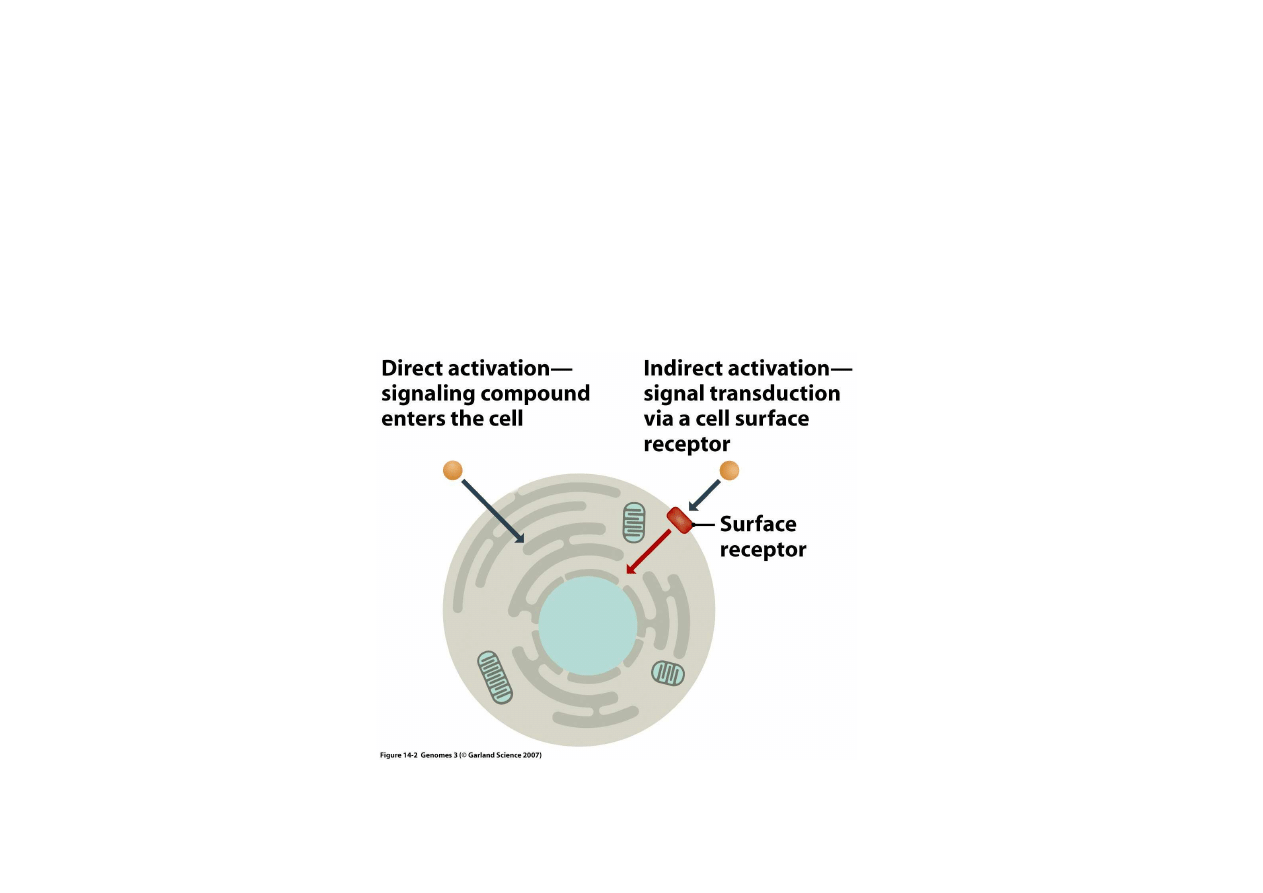
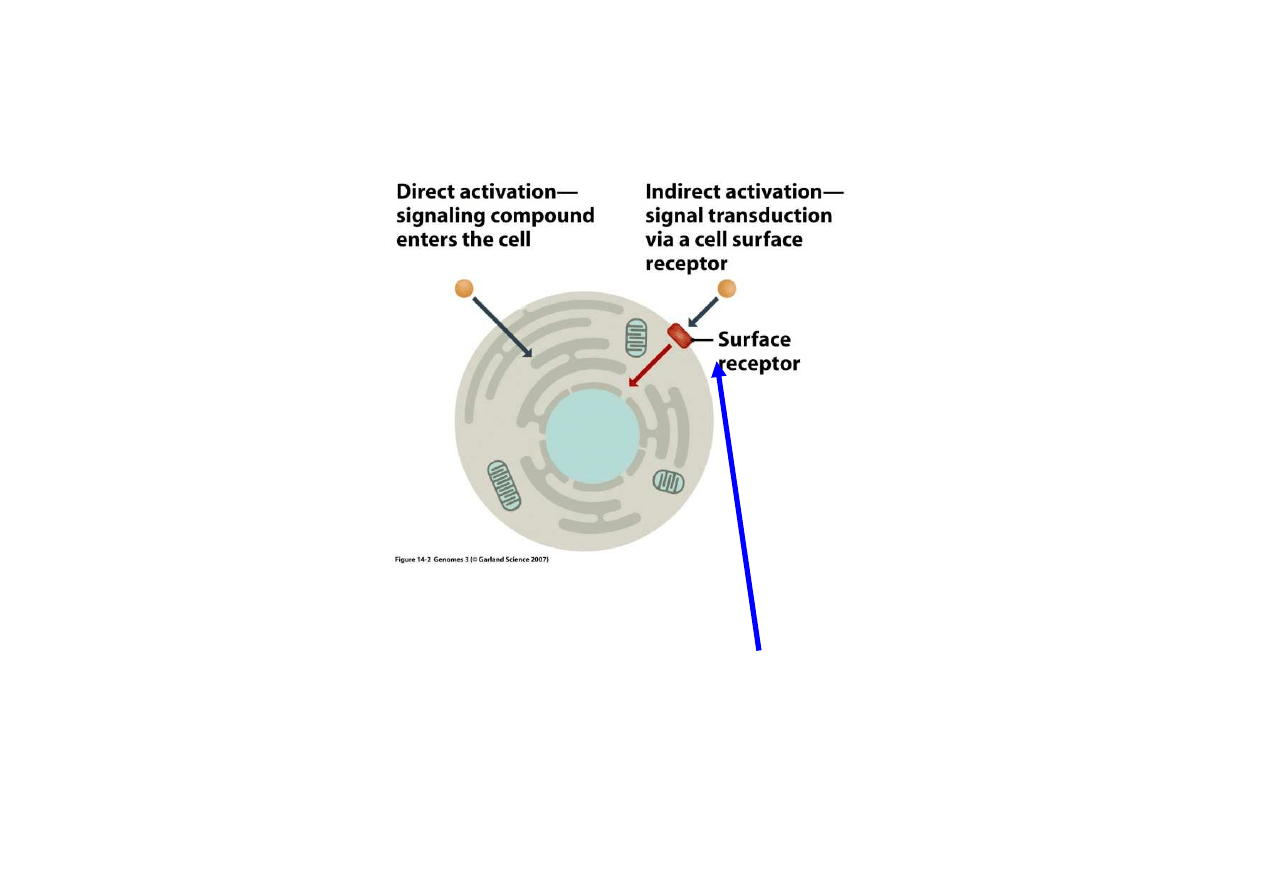
2) Regulacja bezpo
ś
rednia przez czynniki zewn
ę
trzne
Regulacja przez czynniki zewn
ę
trzne odbywa si
ę
na dwa sposoby:
Regulacja po
ś
rednia:
czynnik zewn
ę
trzny
zatrzymuje si
ę
na
powierzchni i
uruchamia czynnik
wewn
ą
trzkomórkowy
Regulacja
bezpo
ś
rednia:
czynnik wnika
do komórki
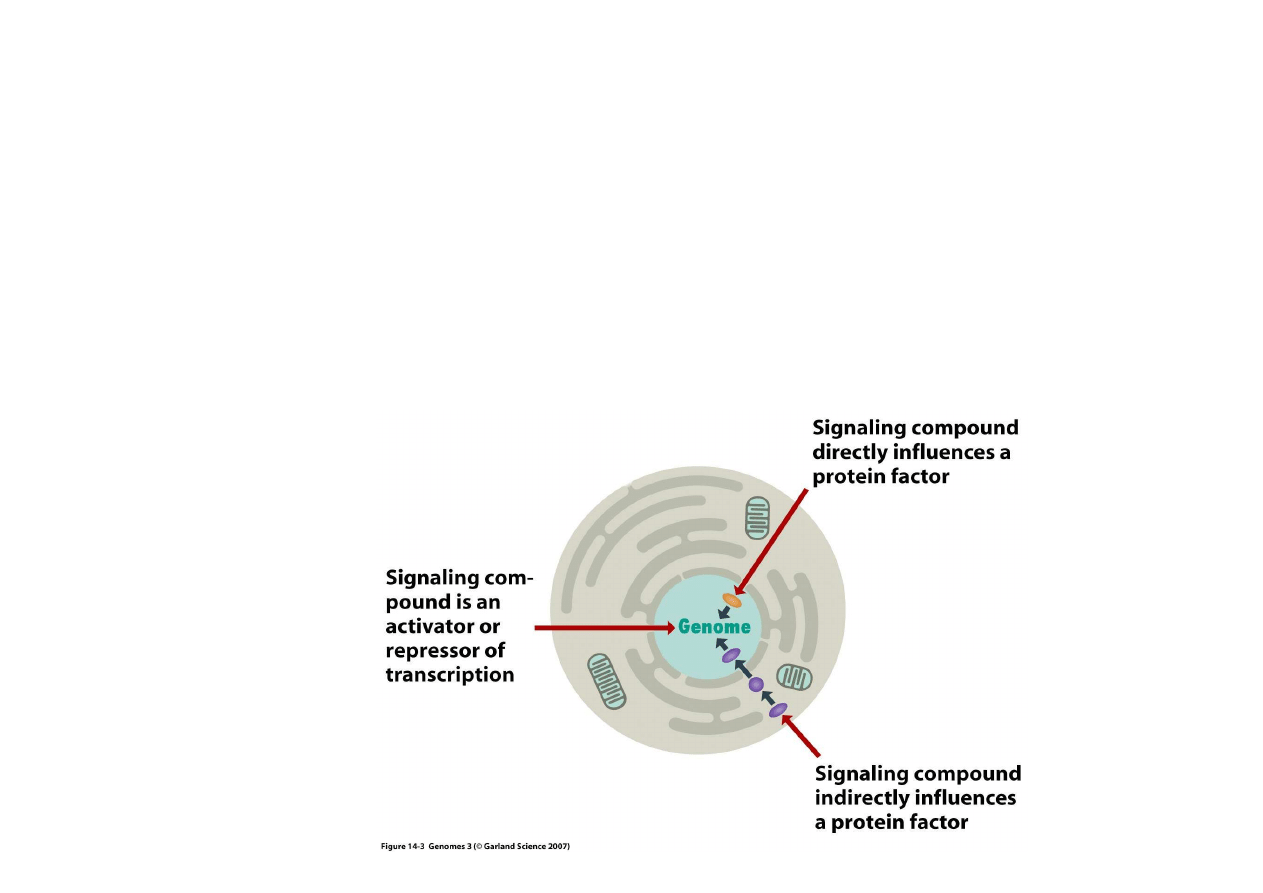
2) Regulacja bezpo
ś
rednia przez czynniki zewn
ę
trzne
Czynnikami sygnałowymi, które wnikaj
ą
bezpo
ś
rednio do komorki mog
ą
by
ć
:
- białka (jak np. laktoferyna), mog
ą
wtedy działa
ć
jakby same były czynnikami
reguluj
ą
cymi transkrypcj
ę
- mniejszymi molekułami (jak np. atomy metali lub cz
ą
steczki hormonów),
wpływaj
ą
cymi na białka regulatorowe
- czynnikami wpływaj
ą
cymi na białka po
ś
rednie, które potem ł
ą
cz
ą
si
ę
z
czynnikami transkrypcyjnymi
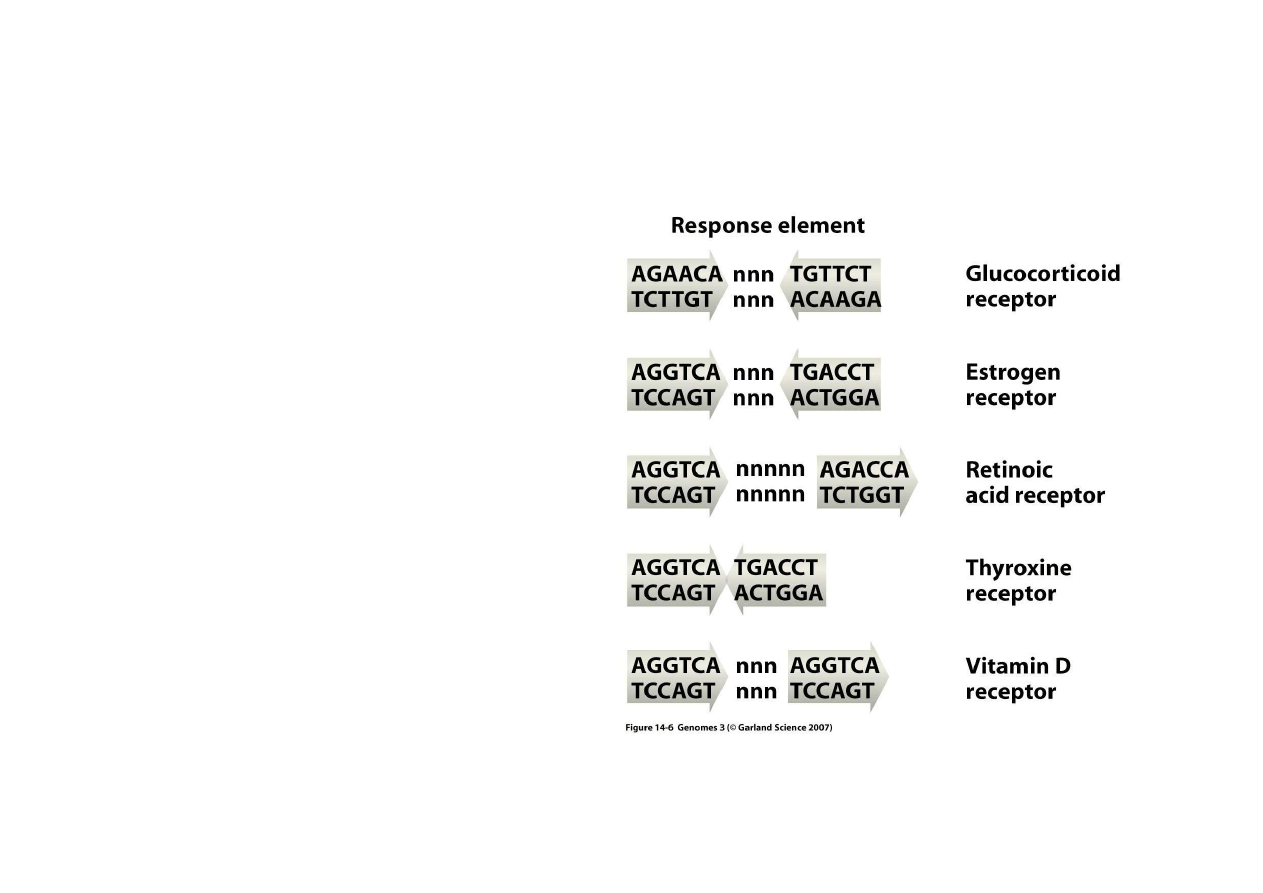
2) Regulacja bezpo
ś
rednia przez czynniki zewn
ę
trzne
Regulacja przez hormony:
Hormony sterydowe ł
ą
cz
ą
si
ę
w
cytoplazmie z białkowymi
receptorami steroidów.
Takie kompleksy w
ę
druj
ą
do
j
ą
dra gdzie ł
ą
cz
ą
si
ę
z DNA w
rejonach przed okre
ś
lon
ą
dla
danego hormonu sekwencj
ą
koduj
ą
c
ą
, co powoduje zmiany
ekspresji genu.
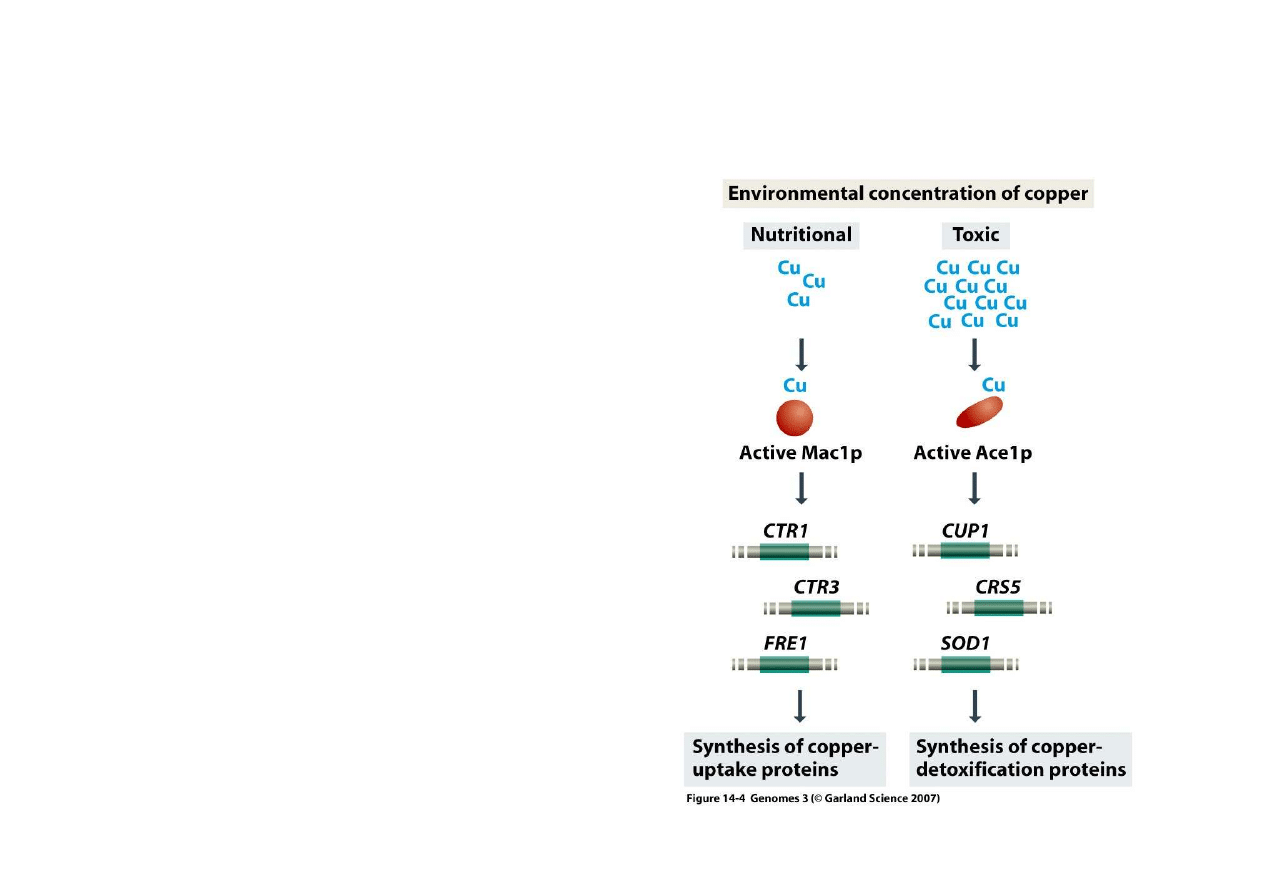
2) Regulacja bezpo
ś
rednia przez czynniki zewn
ę
trzne
Regulacja przez jony:
Ten sam czynnik sygnałowy
mo
ż
e doprowadzi
ć
do ró
ż
nych
skutków, w zale
ż
no
ś
ci od
stanu komórki:
stanu komórki:
- u dro
ż
d
ż
y atom miedzi mo
ż
e
si
ę
poł
ą
czy
ć
z białkiem Mac1 i
(dla pobrania miedzi gdy jej
brakuje)
- z białkiem Ace1 (dla
zmniejszenia toksycznego
efektu miedzi)
3) Regulacja po
ś
rednia poprzez czynniki zewn
ę
trzne
W regulacji po
ś
redniej, czynniki
sygnałowe zewn
ą
trzkomórkowe,
cz
ę
sto natury peptydowej, ł
ą
cz
ą
si
ę
z
receptorami na powierzchni komórki.
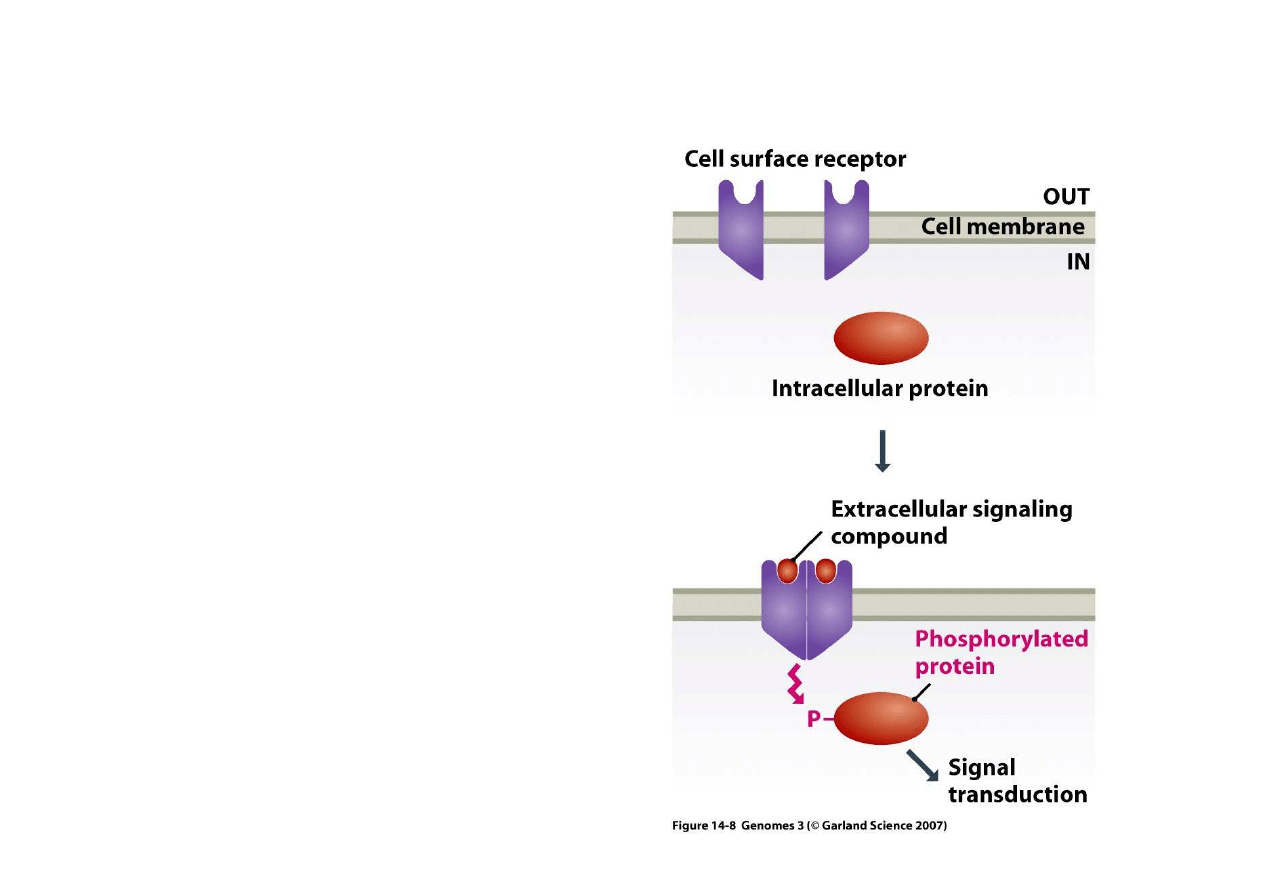
3) Regulacja po
ś
rednia poprzez czynniki zewn
ę
trzne
Poł
ą
czenie czynnika
sygnałowego z
receptorem na
powierzchni komórki
powierzchni komórki
uaktywnia
ś
ci
ś
le okre
ś
lone
białka, bardzo cz
ę
sto
poprzez ich fosforylacj
ę
dokonan
ą
przez
zaktywizowany receptor.
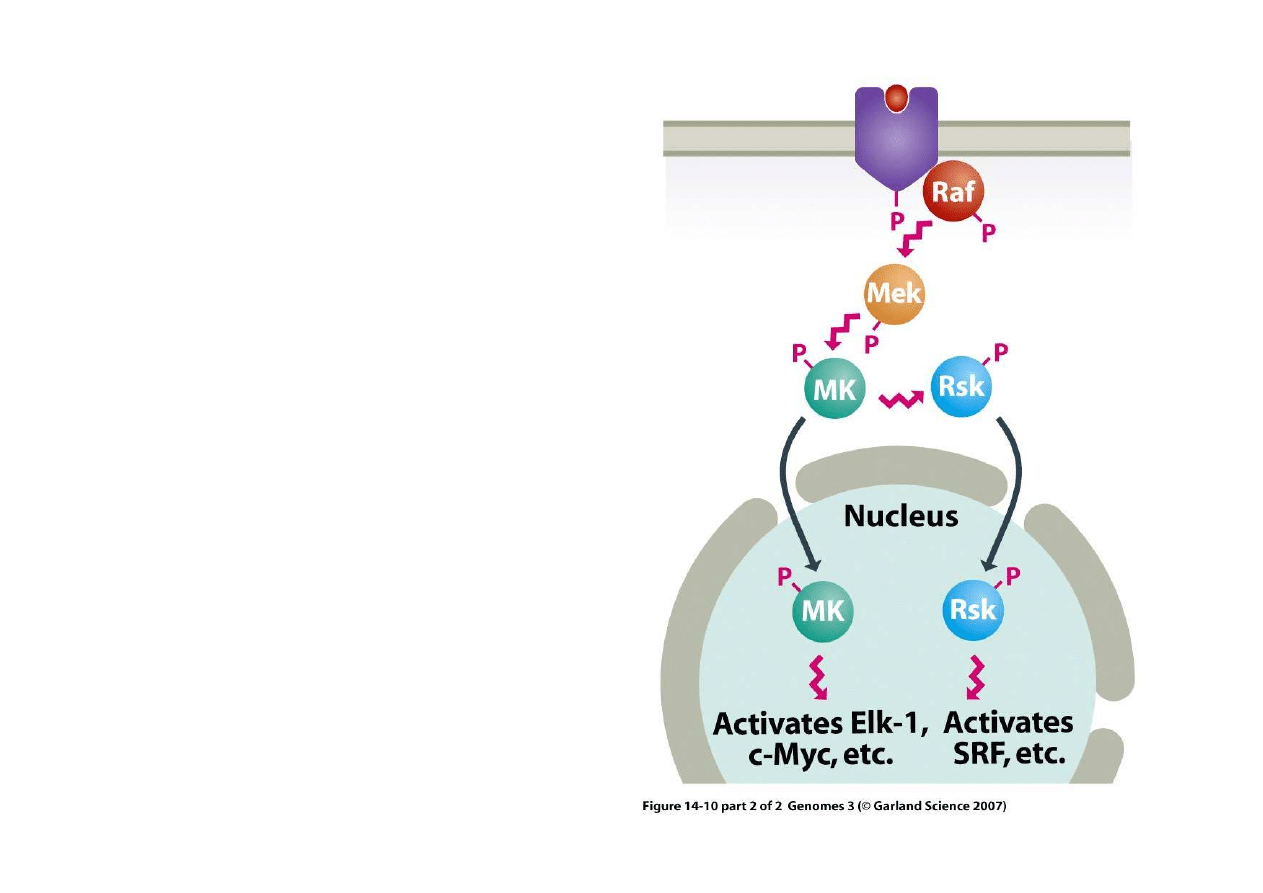
3) Regulacja po
ś
rednia
poprzez czynniki
zewn
ę
trzne
Przykładem działania takiego
sygnału s
ą
szlaki typu MAP
(mitogen activated protein),
Reaguj
ą
one na wiele sygnały
ze
ś
rodowiska i sygnały od
innych komórek (podział).
Sygnał jest wielostopniowy.
Receptor mitogenu (fiolet) po
otrzymaniu sygnału fosforyluje
białko Raf, Raf fosforyluje Mek,
Mek forforyluje MK a MK
fosforyluje albo kolejne białko
po
ś
rednicz
ą
ce (Rsk) albo
ostateczne białkowe czynniki
transkrypcyjne (Elk-1, c-Myc i
inne)
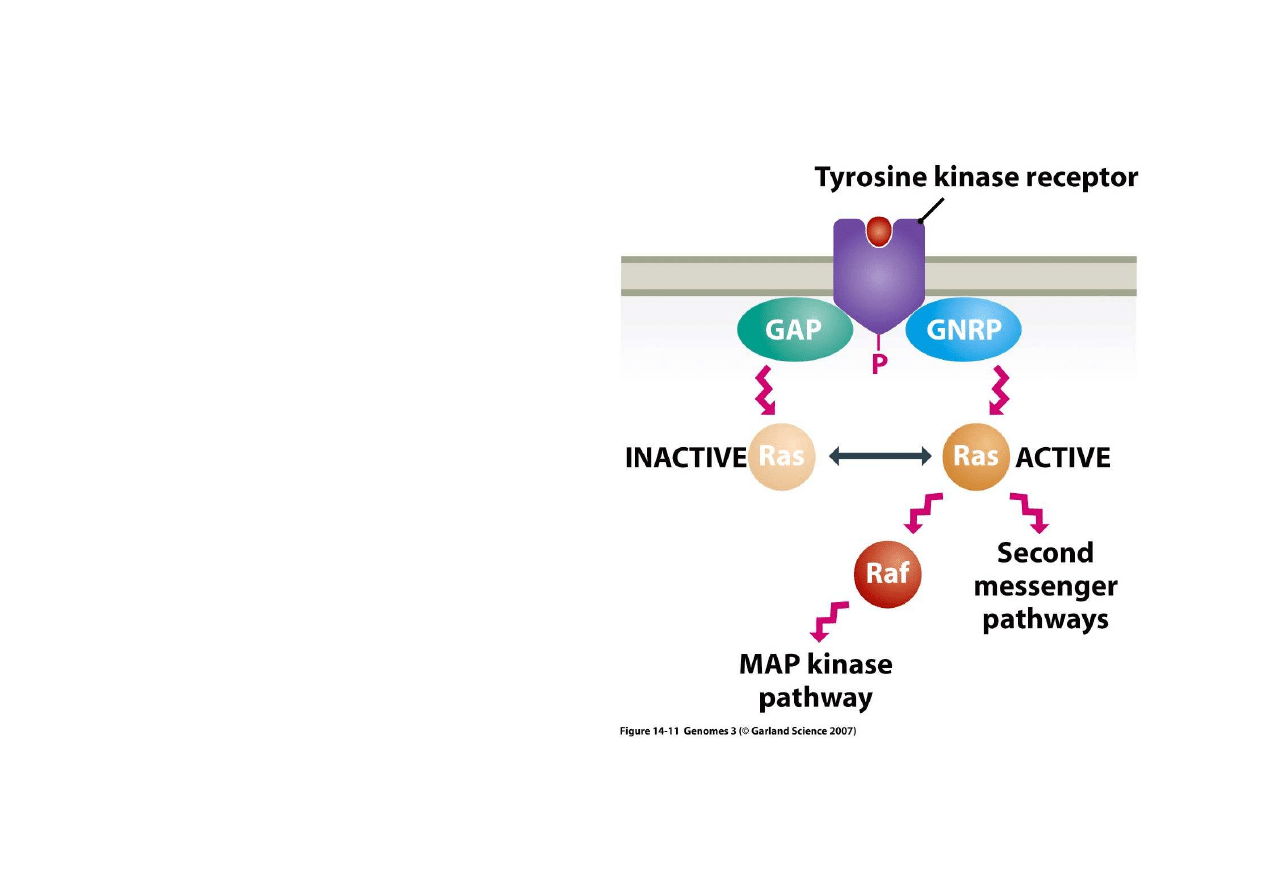
3) Regulacja po
ś
rednia
poprzez czynniki
zewn
ę
trzne
Wielostopniowo
ść
pomaga w
składaniu i rozdzielaniu
sygnałów.
Szlak Ras jest wa
ż
ny w
sygnalizacji wzrostu komórki,
jego deregulacja prowadzi do
nowotworu.
nowotworu.
Powierzchniowa kinaza
tyrozynowa mo
ż
e ł
ą
czy
ć
si
ę
z
białkiem GAP lub GNRP co
odpowiednio inaktywuje lub
aktywuje białko Ras.
Ras mo
ż
e poprzez białko Raf
wpłyn
ąć
na aktywno
ść
szlaku
podziału (MAP) co jest logiczne,
bo wzrost i podział musza by
ć
zsynchronizowane.
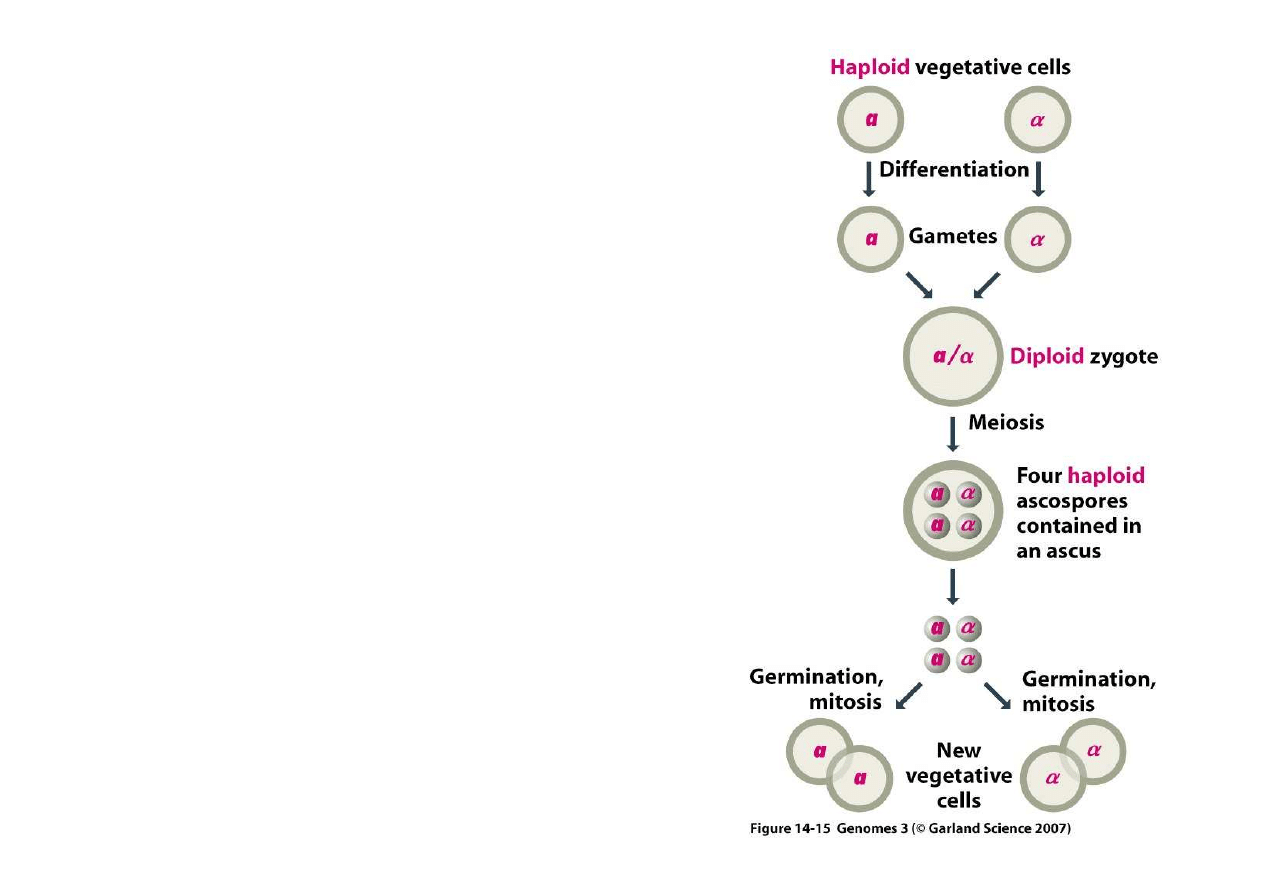
4) Trwałe zmiany regulacyjne poprzez
rearan
ż
acje genów
– typ kojarzeniowy dro
ż
d
ż
y
Dro
ż
d
ż
e produkuj
ą
haploidalne
dro
ż
d
ż
e o okre
ś
lonym typie
kojarzeniowym.
Skojarzenie z haploidem o
przeciwnym typie prowadzi do
odtworzenia diploidalno
ś
ci.
odtworzenia diploidalno
ś
ci.
Gdy komórek o przeciwnym typie
nie ma, haploid mo
ż
e zmieni
ć
typ
przy okazji podziału i skojarzy
ć
si
ę
z komórk
ą
siostrzan
ą
(która ma typ
nie zmieniony)
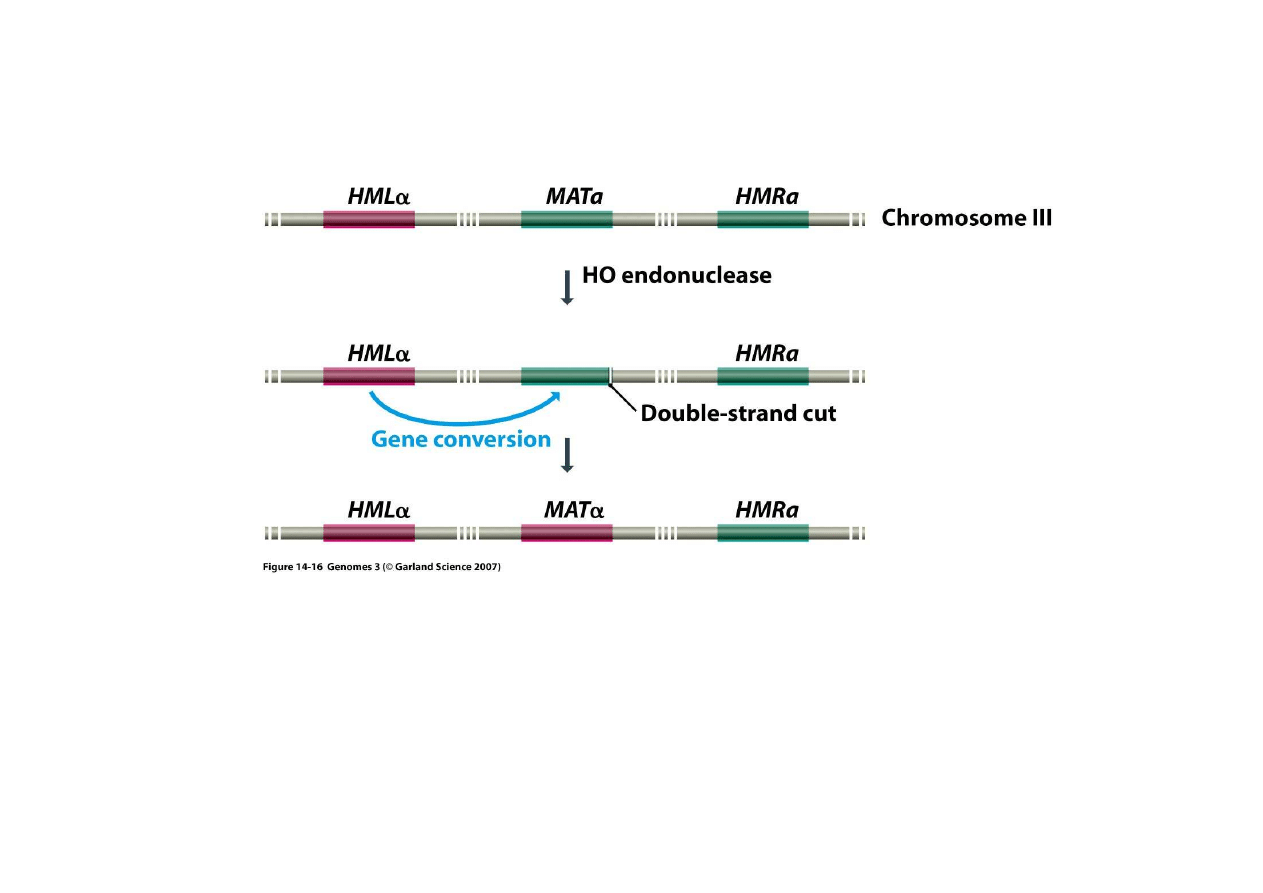
4) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– typ kojarzeniowy dro
ż
d
ż
y
Dro
ż
d
ż
e posiadaj
ą
trzy kopie genu (allele) odpowiadaj
ą
ce za białko
warunkuj
ą
ce typ kojarzeniowy, jakby trzy kasety, ale tylko jeden z nich jest
w locus o czynnej transkrypcji (
ś
rodkowy), tak jakby kaseta w odtwarzaczu.
Wymiana kaset dokonuje si
ę
poprzez zap
ę
tlenie nici DNA i konwersj
ę
genu
na drodze rekombinacji homologicznej.
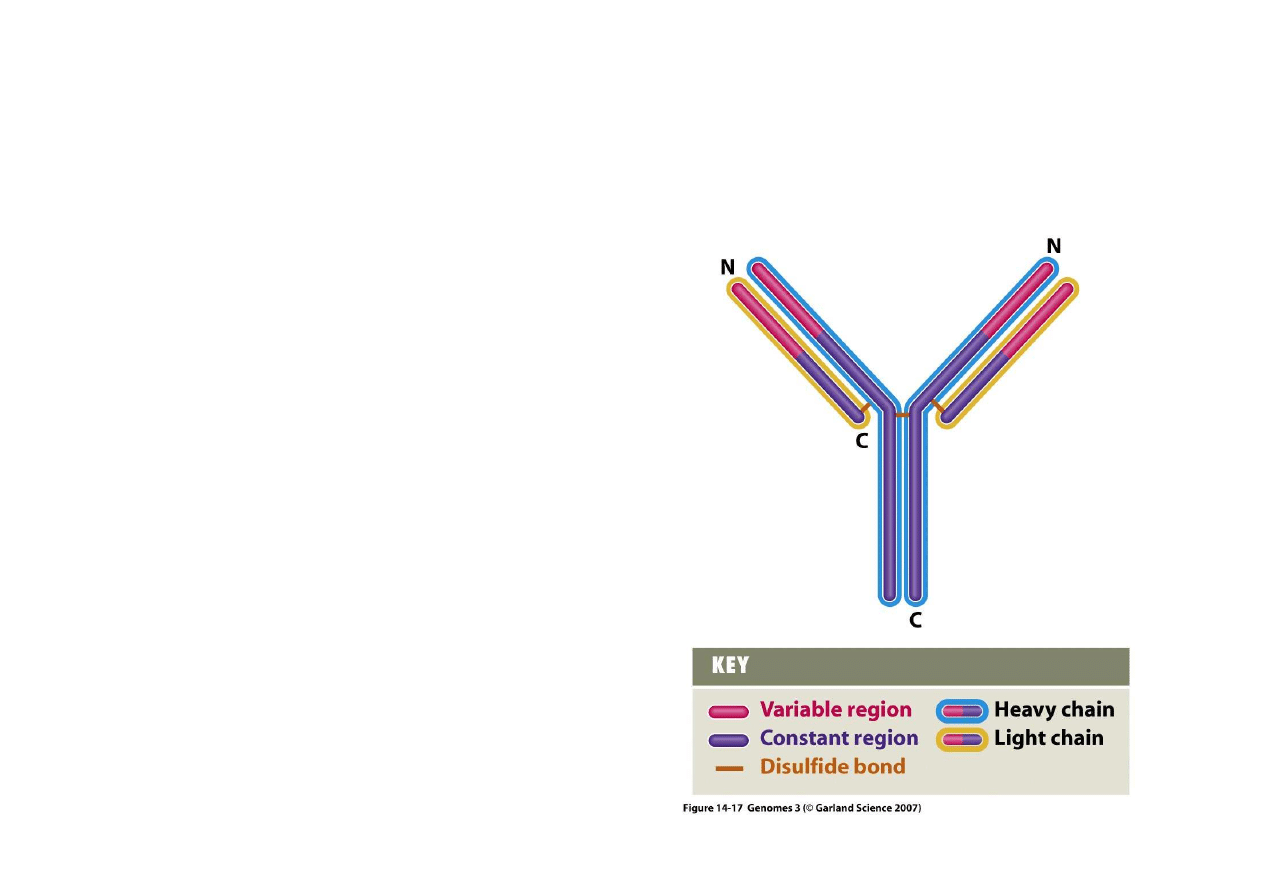
5) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– powstawanie immunoglobulin
Immunoglobuliny ssaków (a tak
ż
e
receptory komórek T i białka
zgodno
ś
ci tkankowej MHC) s
ą
bardzo
ró
ż
ne w ró
ż
nych komórkach tego
samego osobnika.
Ró
ż
norodno
ść
ta powstaje na skutek
rearan
ż
acji, która ma wspólne zasady,
ale ró
ż
ny szczegółowy przebieg w
poszczególnych komórkach.
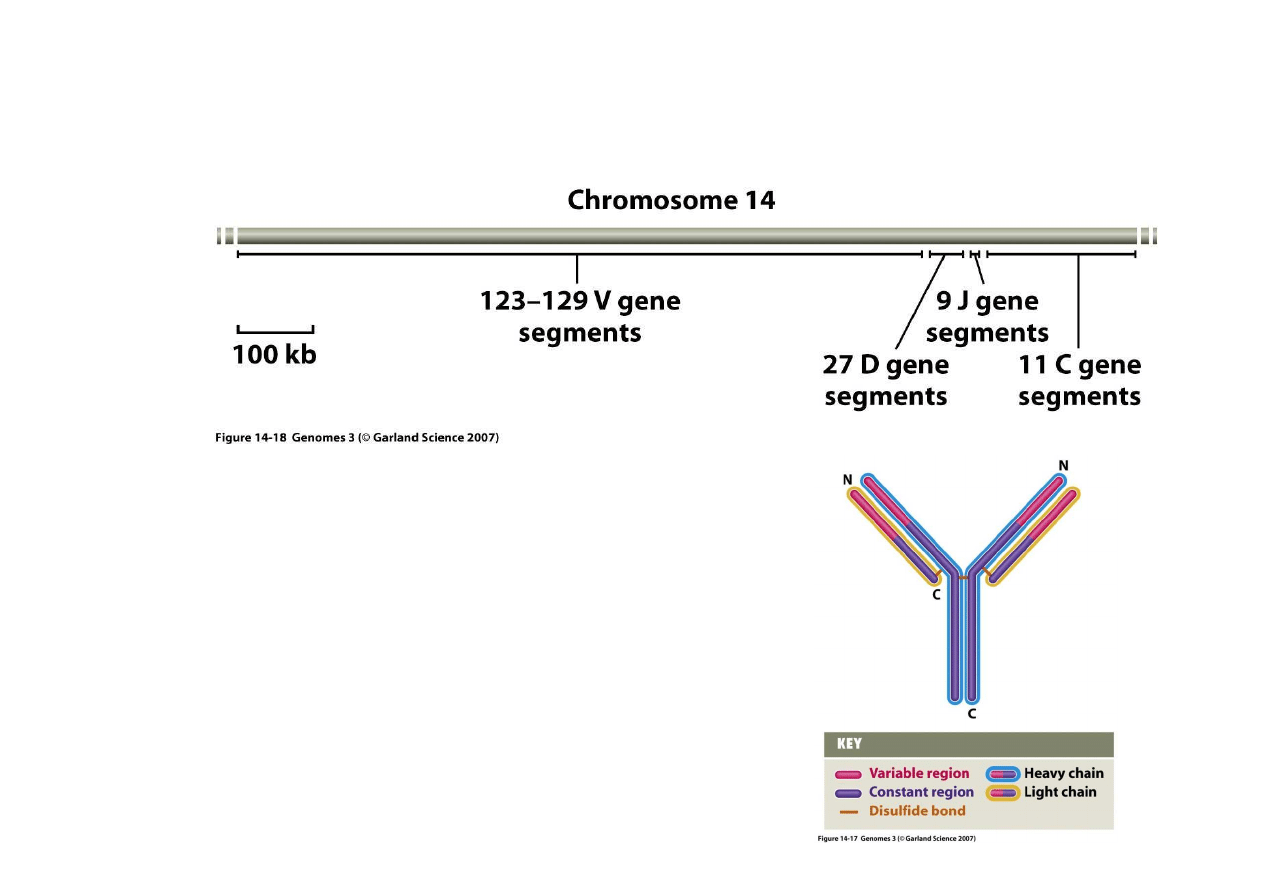
5) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– powstawanie immunoglobulin
Dla ła
ń
cuchów lekkich i ci
ęż
kich
immunoglobulin istniej
ą
osobne rejony
chromosomowe koduj
ą
ce wiele segmentów
cz
ęś
ci zmiennych (V) i po kilka segmentów
cz
ęś
ci ł
ą
cznikowych (D i J) oraz stałych (C).
Powy
ż
ej przykład takiego rejonu koduj
ą
cego
ła
ń
cuch ci
ęż
ki na chromosomie 14 człowieka.
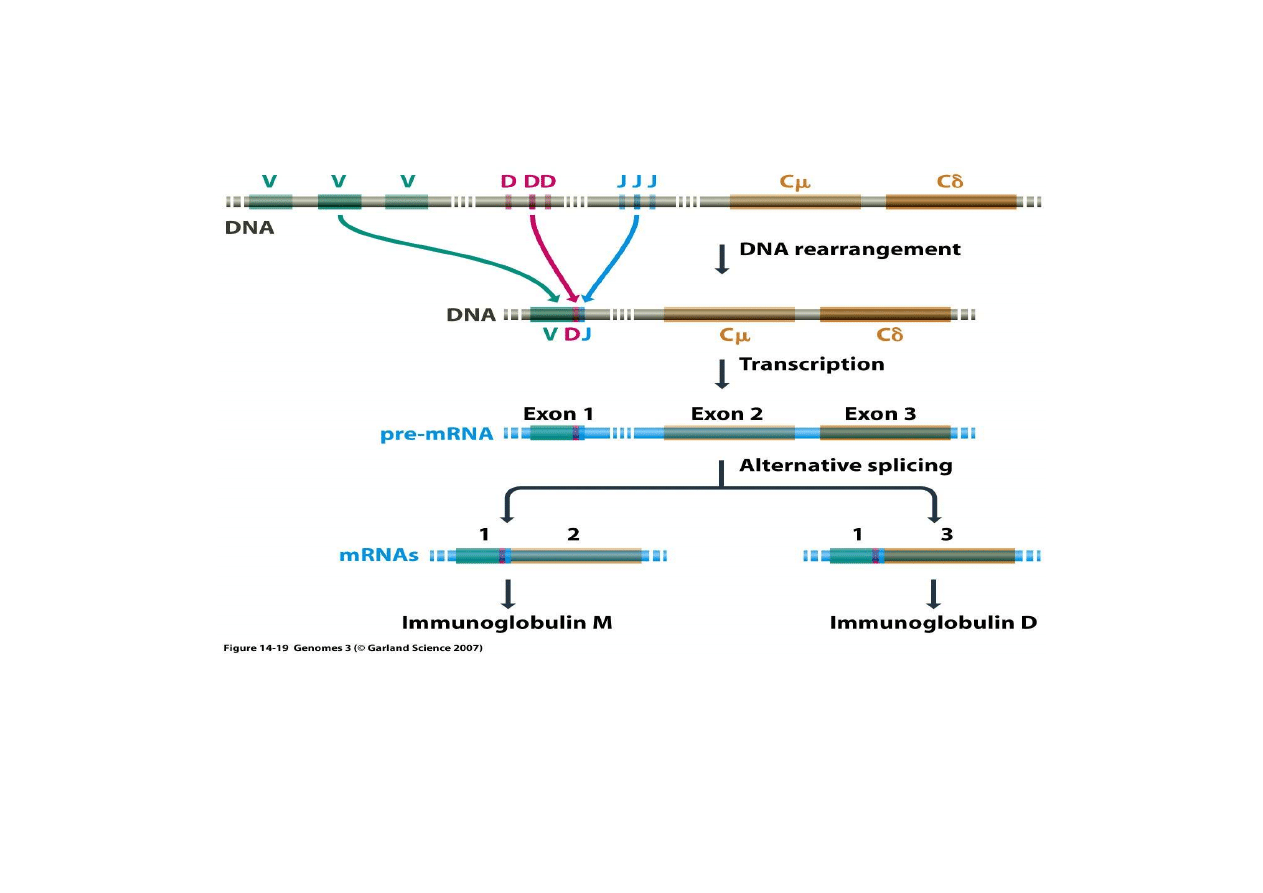
5) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– powstawanie immunoglobulin
W dojrzewaj
ą
cych limfocytach dochodzi do usuni
ę
cia du
ż
ych odcinków DNA (ró
ż
nych
w ró
ż
nych komórkach) tak,
ż
e powstaje tylko jedna kombinacja odcinków VDJ z
dwoma odcinkami C. Alternatywne składanie egzonów doprowadza do wyboru
jednego z tych pocz
ą
tkowo dwóch C.
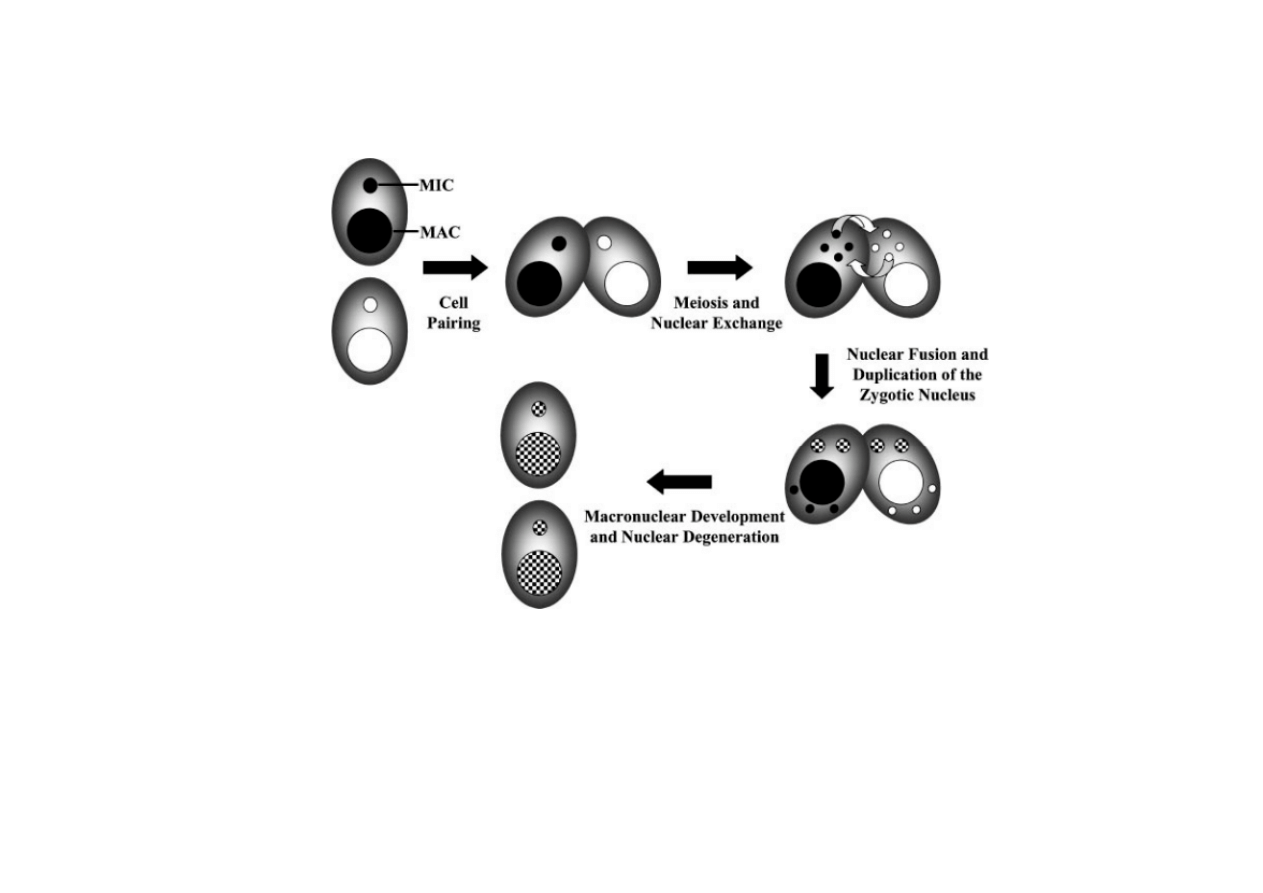
6) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– makronukleus orz
ę
sków
Pantofelek ma mikronukleus (MIC) i makronukleus (MAC), w koniugacji
wymieniane s
ą
haploidalne MIC, one daj
ą
nowe diploidalne MIC, z których
rozwija si
ę
nowy MAC, a stary zanika.
W MIC nie ma transkrypcji, ale jest tam komplet informacji genetycznej.
Transkrypcja jest w MAC, gdzie jest tylko cz
ęść
genów ale za to w wielu
kopiach.
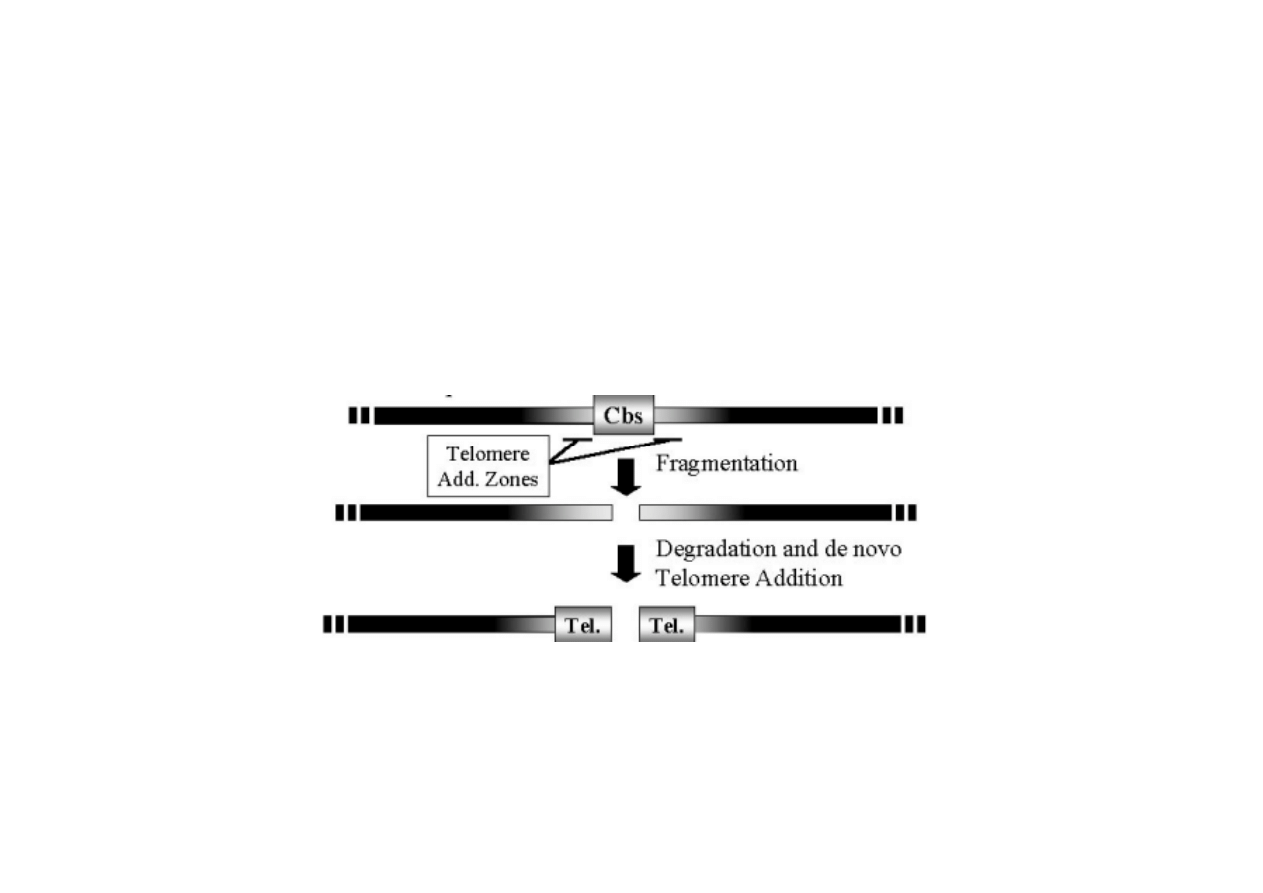
6) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– makronukleus orz
ę
sków
W czasie rozwoju MIC dochodzi do poci
ę
cia chromosomów poł
ą
czonego z
syntez
ą
nowych telomerów lub do składania poci
ę
tych fragmentów, przy
czym ich kolejno
ść
mo
ż
e by
ć
zmieniona
W powy
ż
szym przykładzie, DNA jest ci
ę
te w zakonserwowanych
sekwencjach Cbs a potem z obu stron doczepiane s
ą
telomery
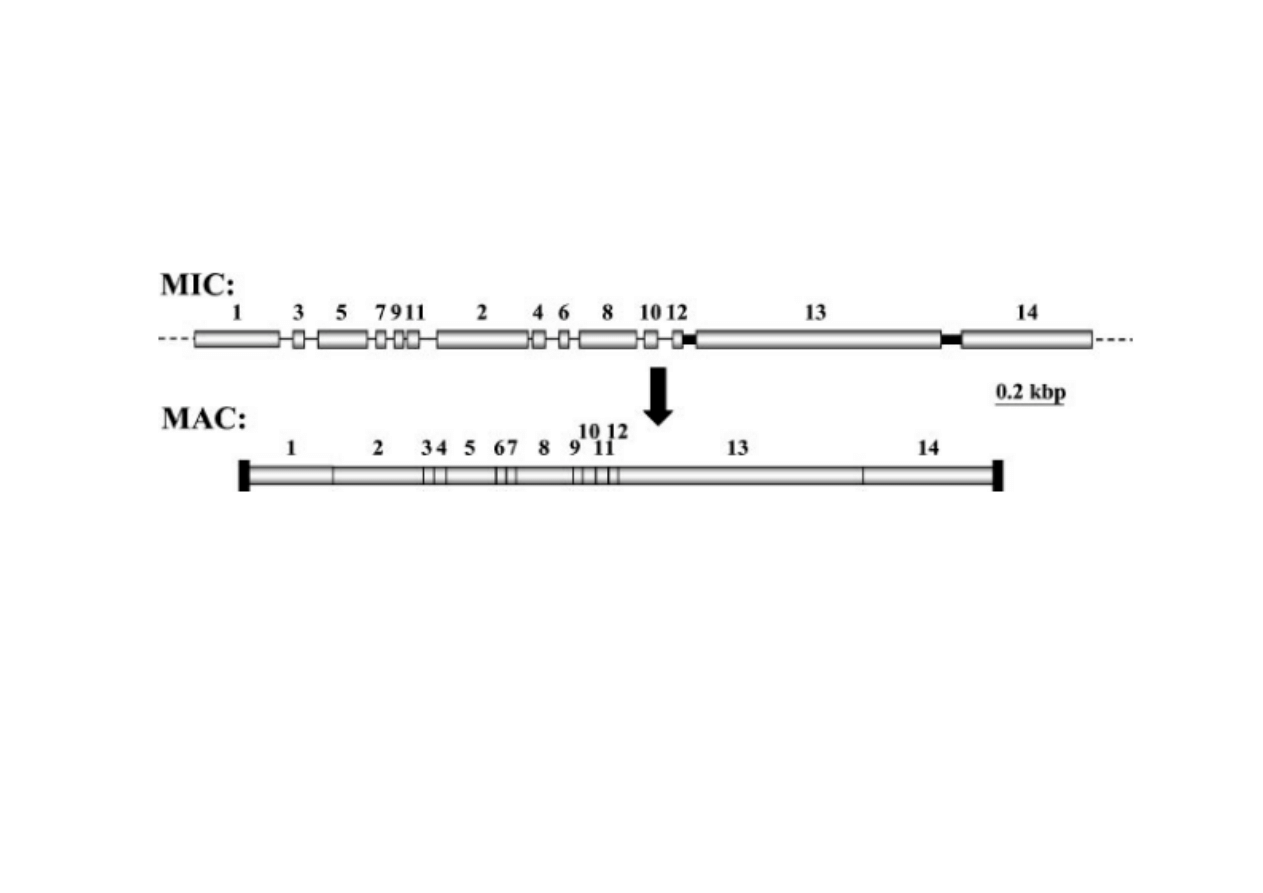
6) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– makronukleus orz
ę
sków
Przykład genu koduj
ą
cego białko doczepione do telomerów. Cz
ęść
sekwencji MIC została utracona, a uzyskane fragmenty maja inn
ą
kolejno
ść
po zło
ż
eniu w MAC. Tylko gen w MAC ma funkcjonaln
ą
sekwencj
ę
. Ocenia si
ę
,
ż
e podczas dojrzewania MAC w całym
genomie wyci
ę
ciu i utraceniu ulega około 60000 fragmentów.

6) Trwałe zmiany regulacyjne poprzez rearan
ż
acje genów
– makronukleus orz
ę
sków
Eliminacja DNA z MIC zaczyna si
ę
od
dwuniciowych naci
ęć
DNA a jej przebieg
jest prawdopodobnie kierowany głównie
poprzez oddziaływanie DNA ze
specjalnie do tego celu syntetyzowanym
niekoduj
ą
cym RNA.
Rearan
ż
acje i zwielokrotnienie kopii
DNA przy syntezie MAC s
ą
ekstremalnym przykładem trwałych
zmian genomu somatycznego, tłumacz
ą
je ogromne rozmiary komórek orz
ę
sków.
Wyszukiwarka
Podobne podstrony:
Regulacja ekspresji genow poprawione
Regulacja ekspresji genów u eucaryota
3. Przyklady regulacji ekspresji genow u Eukaryota-ok, Biologia II, Biologia molekularna
Regulacja ekspresji genów, fizjoterapia, biologia medyczna
1 Regulacja ekspresji genów u prokariota
Regulacja ekspresji genów - materiały teoretyczne
Epigenetyczne mechanizmy regulacji ekspresji genow?
W3 Regulacja ekspresji genów
wd 4 glukoza, lipidy metabolizm, regulatory ekspresji genów
Regulacja ekspresji genów egzamin
Regulacja Ekspresji genow
07) Regulacja ekspresji genów (wyklad 7)
Regulacja ekspresji genów
Regulacja ekspresji genow
Gradient ekspresji genów w regulacji morfogenezy u ssaków, Medycyna ŚUM, Rok 1, Biologia medyczna, T
Gradient ekspresji genów w regulacji morfogenezy u ssaków, Medycyna ŚUM, Rok 1, Biologia medyczna, T
Genetyka regulacja funkcji genow
Ekspresja genów
4 Regulacja ekspresji
więcej podobnych podstron