
Białka biorące
udział w skurczu
mięśnia
Łukasz Bryl gr. 1

Budowa miofibryli - sarkomer

Miozyna
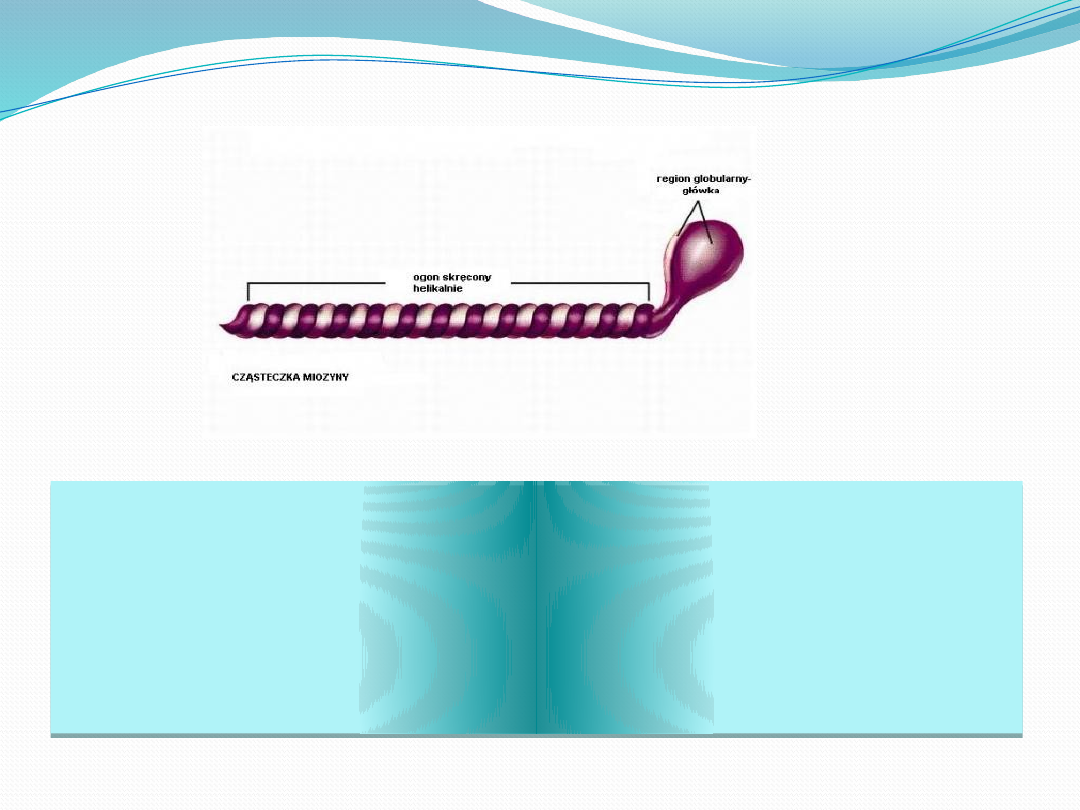
białko o masie 480kDa
Składa się z dwóch łańcuchów ciężkich i dwóch par
łańcuchów lekkich.
N-końcowe części ciężkich łańcuchów zwinięte są w
globularne „główki”, a u ich podstawy zlokalizowane są po
dwa lekkie łańcuchy
Ciężkie łańcuchy owijają się wokół siebie, tworząc długą
superhelisę.
za pomocą główek następuje wiązanie i hydroliza ATP, oraz
wiązanie głównego białka cienkich filamentów - aktyny.
Poszczególne cząsteczki miozyny w filamencie grubym
wiąże ze sobą białko C.
białko o masie 480kDa
Składa się z dwóch łańcuchów ciężkich i dwóch par
łańcuchów lekkich.
N-końcowe części ciężkich łańcuchów zwinięte są w
globularne „główki”, a u ich podstawy zlokalizowane są po
dwa lekkie łańcuchy
Ciężkie łańcuchy owijają się wokół siebie, tworząc długą
superhelisę.
za pomocą główek następuje wiązanie i hydroliza ATP, oraz
wiązanie głównego białka cienkich filamentów - aktyny.
Poszczególne cząsteczki miozyny w filamencie grubym
wiąże ze sobą białko C.
W globularnej głowie miozyny znajdują się
dwa miejsca aktywne:
1.
Służące do enzymatycznego rozpadu ATP
2.
Miejsce wiązania z aktyną
W globularnej głowie miozyny znajdują się
dwa miejsca aktywne:
1.
Służące do enzymatycznego rozpadu ATP
2.
Miejsce wiązania z aktyną

Miozyna
Meromiozyna

Główka miozyny ma domenę regulatorową
i domenę motoryczną
Domena motoryczna dzieli się na 4
subdomeny:
N-końcową
Subdomenę
Subdomenę
Subdomenę


Domena motoryczna ma szczelinę
oddzielającą od siebie 2 subdomeny, w
pobliżu ujścia tej szczeliny jest miejsce
wiązania aktyny
Podstawa szczeliny to podstawa kieszeni
katalitycznej , gdzie wiązany jest ATP lub
produkty hydrolizy ATP.
Domena motoryczna ma szczelinę
oddzielającą od siebie 2 subdomeny, w
pobliżu ujścia tej szczeliny jest miejsce
wiązania aktyny
Podstawa szczeliny to podstawa kieszeni
katalitycznej , gdzie wiązany jest ATP lub
produkty hydrolizy ATP.

Filamenty cienkie
Miofilamenty cienkie stanowią nić białkową, złożoną
z cząsteczek białka globularnego-G-aktyny.
Cząsteczki te w obecności ATP są ze sobą silnie
spolimeryzowane, tworząc sznur zwany F-aktyną.
Każdy miofilament cienki zbudowany jest z dwóch
spiralnie wokół siebie skręconych sznurów F-aktyny.
Oprócz aktyny w skład miofilamentów cienkich
wchodzą białka regulatorowe: troponina i
tropomiozyna
Cienki filament zawiera 1 cząsteczkę tropomiozyny i 1
troponiny na każde 7 monomerów aktyny.
Miofilamenty cienkie stanowią nić białkową, złożoną
z cząsteczek białka globularnego-G-aktyny.
Cząsteczki te w obecności ATP są ze sobą silnie
spolimeryzowane, tworząc sznur zwany F-aktyną.
Każdy miofilament cienki zbudowany jest z dwóch
spiralnie wokół siebie skręconych sznurów F-aktyny.
Oprócz aktyny w skład miofilamentów cienkich
wchodzą białka regulatorowe: troponina i
tropomiozyna
Cienki filament zawiera 1 cząsteczkę tropomiozyny i 1
troponiny na każde 7 monomerów aktyny.

Aktyna
jest głównym składnikiem cienkich filamentów
monomer o masie 43 kDa, zwany aktyną G,
ponieważ ma kształt globularny
aktyna G polimeryzuje do formy fibrylarnej
zwanej aktyną F
podobnie jak miozyna jest ATP-azą. Jednak
hydroliza ATP przez aktynę nie wywołuje
skurczu mięśnia. Cykl ATP-ADP aktyny bierze
udział w polimeryzacji i depolimeryzacji
filamentu.
jest głównym składnikiem cienkich filamentów
monomer o masie 43 kDa, zwany aktyną G,
ponieważ ma kształt globularny
aktyna G polimeryzuje do formy fibrylarnej
zwanej aktyną F
podobnie jak miozyna jest ATP-azą. Jednak
hydroliza ATP przez aktynę nie wywołuje
skurczu mięśnia. Cykl ATP-ADP aktyny bierze
udział w polimeryzacji i depolimeryzacji
filamentu.

Tropomiozyna
Tropomiozyna występuje we
wszystkich rodzajach mięśni jak
również towarzyszy aktynie w
komórkach niemięśniowych.
Cząsteczka Tropomiozyny
zbudowana jest z dwóch łańcuchów
polipeptydowych o strukturze alfa-
helisy skręconych w superhelisę.
W mięśniach poprzecznie
prążkowanych występują dwie
izoformy alfa i beta, które tworzą
homo(np. alfa-alfa) i heterodimery.
Proporcje izoform zależą od rodzaju
mięśnia .
Tropomiozyna występuje we
wszystkich rodzajach mięśni jak
również towarzyszy aktynie w
komórkach niemięśniowych.
Cząsteczka Tropomiozyny
zbudowana jest z dwóch łańcuchów
polipeptydowych o strukturze alfa-
helisy skręconych w superhelisę.
W mięśniach poprzecznie
prążkowanych występują dwie
izoformy alfa i beta, które tworzą
homo(np. alfa-alfa) i heterodimery.
Proporcje izoform zależą od rodzaju
mięśnia .

Cały łańcuch Tm zbudowany jest z wielokrotnie
powtórzonej heptady , czyli 7 aimnokwasów.
Łańcuchy tropomiozyny przebiegają równolegle do nitek
aktynowych i zlokalizowane są w rowkach nici F-aktyny.
Zawiera 7 grup Ala powodujących lokalne zaburzenia
symetrii klasyczne superhelisy i jej nieznaczne
odkształcenie – 7 potencjalnych miejsc wiązania aktyny.
Udział każdego z tych miejsc nie jest równocenny.
Odpowiada za blokowanie miejsca łączenia aktyny z
miozyną
Cały łańcuch Tm zbudowany jest z wielokrotnie
powtórzonej heptady , czyli 7 aimnokwasów.
Łańcuchy tropomiozyny przebiegają równolegle do nitek
aktynowych i zlokalizowane są w rowkach nici F-aktyny.
Zawiera 7 grup Ala powodujących lokalne zaburzenia
symetrii klasyczne superhelisy i jej nieznaczne
odkształcenie – 7 potencjalnych miejsc wiązania aktyny.
Udział każdego z tych miejsc nie jest równocenny.
Odpowiada za blokowanie miejsca łączenia aktyny z
miozyną

Troponina
Domena Globularna
100 C- końcowych aminokwasów
100 C- końcowych aminokwasów
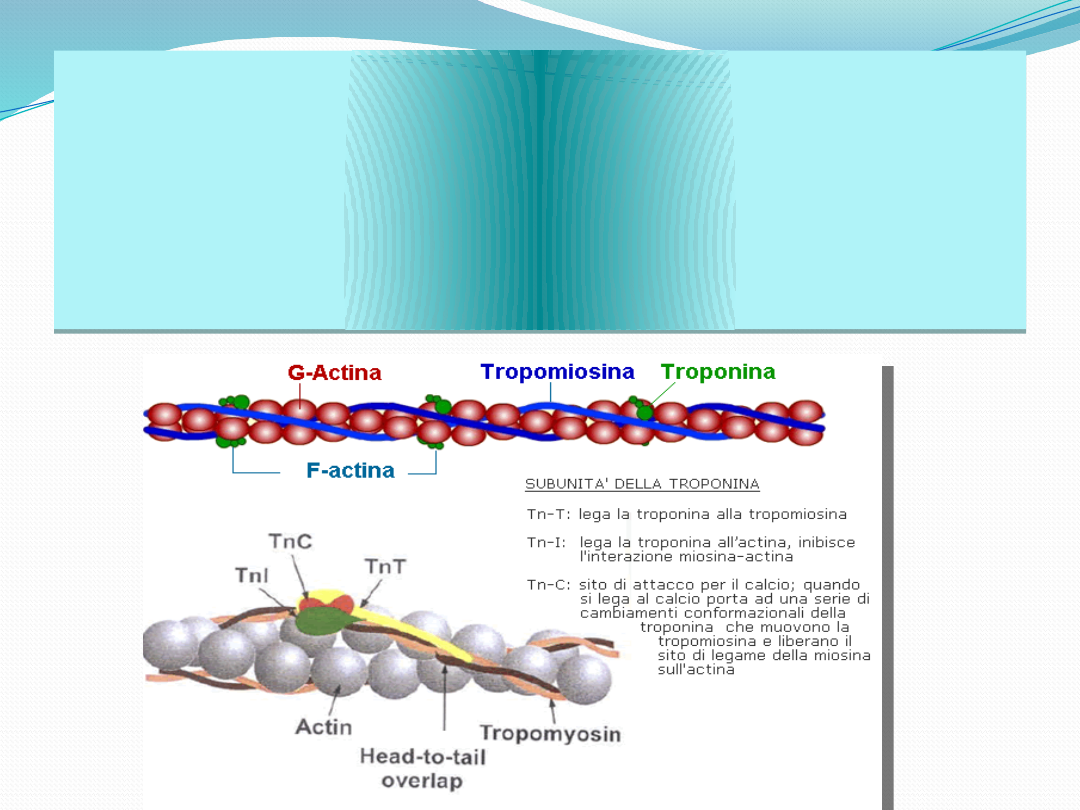
Troponina jest kompleksem trzech białek z których
każde pełni specyficzną rolę w regulacji skurczu
mięśnia . Troponina I hamuje ATPazę
aktomiozynową, troponina C wiąże jony wapnia
Ca
2+
, natomiast troponina T łączy wszystkie
składniki z Tropomiozyną.
Troponina jest kompleksem trzech białek z których
każde pełni specyficzną rolę w regulacji skurczu
mięśnia . Troponina I hamuje ATPazę
aktomiozynową, troponina C wiąże jony wapnia
Ca
2+
, natomiast troponina T łączy wszystkie
składniki z Tropomiozyną.

Troponina I
Tn I wiąże się bezpośrednią z aktyną i hamuje
aktywność ATPazy aktomiozynowej.
Podstawowa rola TnI – utrzymywanie Tm na
powierzchni filamentu aktyny w tzw. „pozycji
blokującej” wiązanie główek miozyny w nieobecności
jonów Ca
2+
.
Tn I zawiera dwa miejsca wiązania TnC. Aminokwasy
będące N- końcem tworzą alfa-helisę, która wiąże się z
hydrofobową kieszenią w C – końcowej domenie TnC.
Wiązanie to wymaga obecności Ca
2+
lub Mg
2+
.
Tn I wiąże się bezpośrednią z aktyną i hamuje
aktywność ATPazy aktomiozynowej.
Podstawowa rola TnI – utrzymywanie Tm na
powierzchni filamentu aktyny w tzw. „pozycji
blokującej” wiązanie główek miozyny w nieobecności
jonów Ca
2+
.
Tn I zawiera dwa miejsca wiązania TnC. Aminokwasy
będące N- końcem tworzą alfa-helisę, która wiąże się z
hydrofobową kieszenią w C – końcowej domenie TnC.
Wiązanie to wymaga obecności Ca
2+
lub Mg
2+
.

Troponina T
Największa podjednostka troponiny.
Podstawową funkcją jest wiązanie Tm fragmentem N-
końcowym TnT.
Alfa helikalny fragment Troponiny T łączy się również
z Troponiną I.
TnT degraduje pod wpływem enzymów tworząc 2
fragmenty: T1 i T2.
T2 tworzy trwały kompleks z TnI i TnC.
Największa podjednostka troponiny.
Podstawową funkcją jest wiązanie Tm fragmentem N-
końcowym TnT.
Alfa helikalny fragment Troponiny T łączy się również
z Troponiną I.
TnT degraduje pod wpływem enzymów tworząc 2
fragmenty: T1 i T2.
T2 tworzy trwały kompleks z TnI i TnC.

Troponina C
Odgrywa kluczową rolę w skurczu.
Posiada zdolność specyficznego wiązania
jonów wapnia w zakresie ich stężeń
molowych.
W TnC występuje charakterystyczny motyw
umożliwiający wiązanie jonów wapnia, helisa-
pętla-helisa (EF-hand) – 2 motywy EF-hand
tworzą domenę strukturalną.
TnC zawiera 2 domeny:
C-końcowa – rola strukturalna
N-końcowa – odpowiada za regulację skurczu
Odgrywa kluczową rolę w skurczu.
Posiada zdolność specyficznego wiązania
jonów wapnia w zakresie ich stężeń
molowych.
W TnC występuje charakterystyczny motyw
umożliwiający wiązanie jonów wapnia, helisa-
pętla-helisa (EF-hand) – 2 motywy EF-hand
tworzą domenę strukturalną.
TnC zawiera 2 domeny:
C-końcowa – rola strukturalna
N-końcowa – odpowiada za regulację skurczu
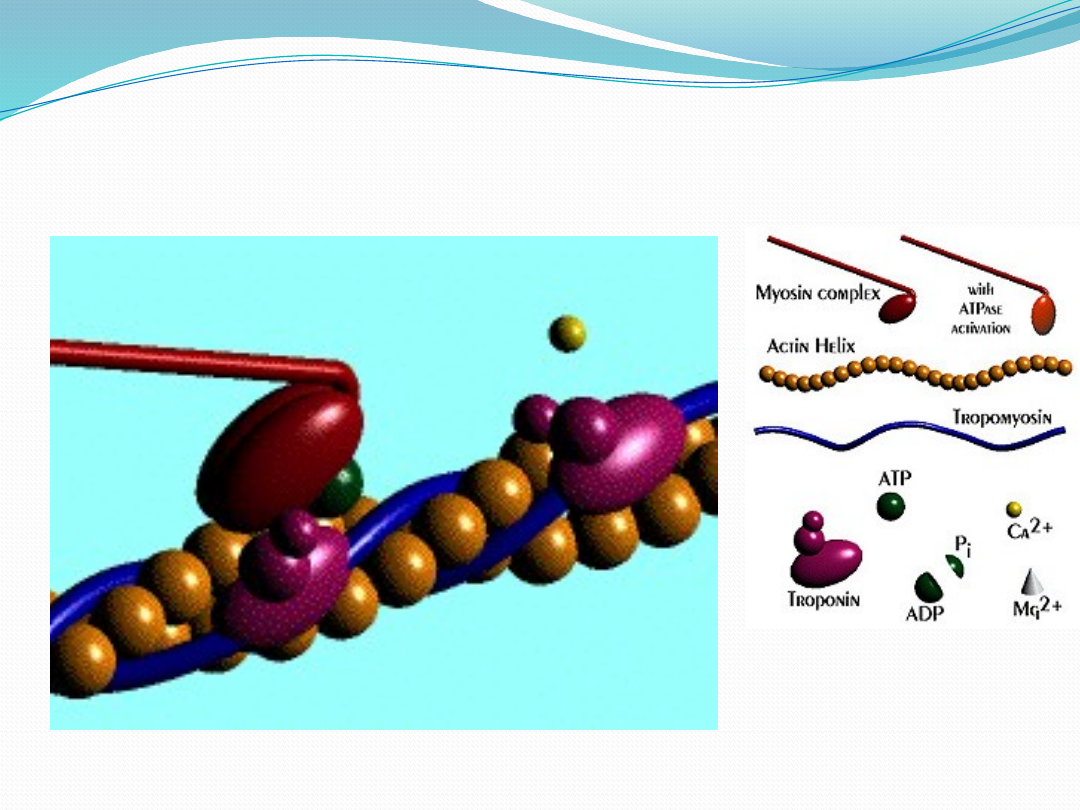
Skurcz mięśnia
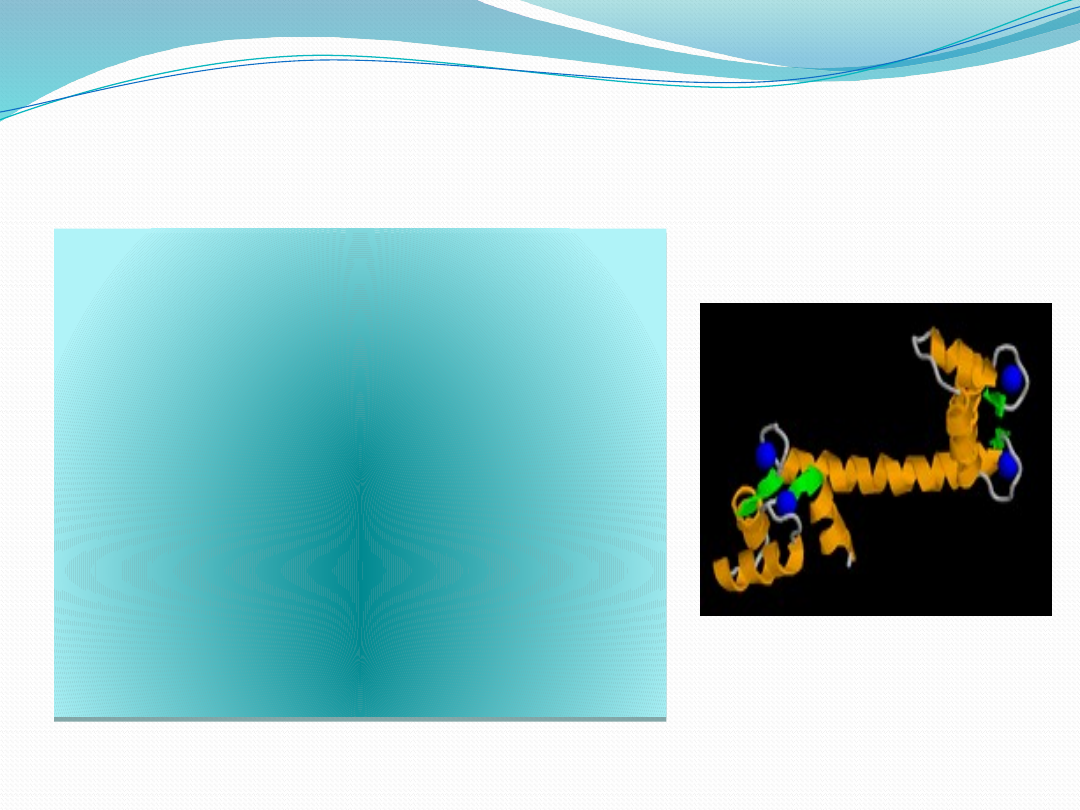
Kalmodulina
Masa cząsteczkowa
kalmoduliny wynosi 16,7
kDa. Jest ona peptydem o
długości 148 aminokwasów.
Jony Ca
2+
łącząc się z
kalmoduliną aktywują kinazę
łańcuchów lekkich miozyny i
prowadzą do fosforylacji tych
łańcuchów, co umożliwia
interakcję aktyny z miozyną
Masa cząsteczkowa
kalmoduliny wynosi 16,7
kDa. Jest ona peptydem o
długości 148 aminokwasów.
Jony Ca
2+
łącząc się z
kalmoduliną aktywują kinazę
łańcuchów lekkich miozyny i
prowadzą do fosforylacji tych
łańcuchów, co umożliwia
interakcję aktyny z miozyną

Kaldesmon(87 kDa)
Cząsteczki kaldesmonu o długości około 75nm,
podobnie jak polimery tropomiozyny, ułożone są
wzdłuż podwójnej helisy monomerów aktyny, w
kontakcie z zewnętrznymi domenami jej cząsteczek.
Jest białkiem zależnym od Ca
2+
, które odgrywa rolę w
regulacji skurczu mięśnia gładkiego. Przy niskich
stężeniach Ca
2+
wiąże się ono z tropomiozyną i aktyną.
Zapobiega to wiązaniu aktyny z miozyną, utrzymując
mięsień w stanie rozkurczu.
w przypadku wyższych stężeń jonów wapnia Ca
2+
-
kalmodulina wiąże kaldesmon, uwalniając go od aktyny
Aktyna wiąże się z miozyną – może zajść skurcz
Cząsteczki kaldesmonu o długości około 75nm,
podobnie jak polimery tropomiozyny, ułożone są
wzdłuż podwójnej helisy monomerów aktyny, w
kontakcie z zewnętrznymi domenami jej cząsteczek.
Jest białkiem zależnym od Ca
2+
, które odgrywa rolę w
regulacji skurczu mięśnia gładkiego. Przy niskich
stężeniach Ca
2+
wiąże się ono z tropomiozyną i aktyną.
Zapobiega to wiązaniu aktyny z miozyną, utrzymując
mięsień w stanie rozkurczu.
w przypadku wyższych stężeń jonów wapnia Ca
2+
-
kalmodulina wiąże kaldesmon, uwalniając go od aktyny
Aktyna wiąże się z miozyną – może zajść skurcz

Inne białka regulujące skurcz mięsni
Białko
Umiejscownienie
Funkcja
Tytyna
Rozciąga się od lini Z do
lini M
Bierze udział w
rozkurczu mięśnia
Nebulina
Od lini Z wzdłuż nitek
aktynowych
Reguluje tworzenie i
długość nitek aktyny
α-aktynina
Zakotwicza aktynę w
liniach Z
Stabilizuje nitki aktyny
Dystrofina
Połączona z plazmalemą
Brak lub niedobór
powoduje upośledzenie
skurczu i rozkurczu
mięśnia
Kalcyneuryna
Cytozol
Odgrywa rolę w
przeroście mięśnia
sercowego oraz w
regulacji liczby szybko i
wolno kurczących się
włókien mięśniowych

Kalcyneuryna
Jest to fosfataza regulowana
przez kalmodulinę. Odgrywa rolę
w regulacji liczby szybko i wolno
kurczących się włókienek
mięśniowych.
Białko tworzy heterodimery,
zbudowane z podjednostki
katalitycznej (kalcyneuryna A) i
regulatorowej (kalcyneuryna B).
Podjednostka B zawiera 4
miejsca wiążące jony wapnia.
Jest to fosfataza regulowana
przez kalmodulinę. Odgrywa rolę
w regulacji liczby szybko i wolno
kurczących się włókienek
mięśniowych.
Białko tworzy heterodimery,
zbudowane z podjednostki
katalitycznej (kalcyneuryna A) i
regulatorowej (kalcyneuryna B).
Podjednostka B zawiera 4
miejsca wiążące jony wapnia.

Tytyna
długie, włókniste białko mięśni poprzecznie
prążkowanych masie cząsteczkowej około 3x10
6
Da,
Biegnie przez pół sarkomeru łącząc prążki Z i M
Część łańcucha przebiegająca w prążku A posiada
okresowo powtarzające się domeny wiążące się z
lekka meromiozyną.
Tytyna jest pofałdowana tworząc rodzaj sprężyny,
która zostaje ściśnięta w czasie skurczu i
rozprężona w czasie rozkurczu. Białko te jest
elementem sprężystym sarkomeru.
długie, włókniste białko mięśni poprzecznie
prążkowanych masie cząsteczkowej około 3x10
6
Da,
Biegnie przez pół sarkomeru łącząc prążki Z i M
Część łańcucha przebiegająca w prążku A posiada
okresowo powtarzające się domeny wiążące się z
lekka meromiozyną.
Tytyna jest pofałdowana tworząc rodzaj sprężyny,
która zostaje ściśnięta w czasie skurczu i
rozprężona w czasie rozkurczu. Białko te jest
elementem sprężystym sarkomeru.

Nebulina
Jest białkiem fibrylarnym o długości równej dł.
cienkiej nitki sarkomeru, wokół którego jest
owinięte.
Posiada okresowo powtarzające się domeny
wiążące aktynę. Swoim końcem jest
zakotwiczona w prążku Z.
Utrzymuje ułożenie filamentów cienkich w
sarkomerze.
Jest białkiem fibrylarnym o długości równej dł.
cienkiej nitki sarkomeru, wokół którego jest
owinięte.
Posiada okresowo powtarzające się domeny
wiążące aktynę. Swoim końcem jest
zakotwiczona w prążku Z.
Utrzymuje ułożenie filamentów cienkich w
sarkomerze.

Desmina
Tworzy filamenty pośrednie typu III, które
łączą ze sobą linie Z sąsiednich miofibryli
Tworzy filamenty pośrednie typu III, które
łączą ze sobą linie Z sąsiednich miofibryli
α-aktynina
α-aktynina – zakotwicza aktynę w liniach Z
Stabilizuje nitki aktyny
α-aktynina – zakotwicza aktynę w liniach Z
Stabilizuje nitki aktyny
Białko c
wiąże miozynę i tytnę odgrywa rolę w
integracji struktury sarkomeru
wiąże miozynę i tytnę odgrywa rolę w
integracji struktury sarkomeru

Dystrfofina
Dystrofina składa się z 3684 aminokwasów i ma masę 427 kDa.
Dystrofina jest białkiem kodowanym przez gen leżący na chromosomie X.
Rola dystrofiny polega na "uszczelnianiu" błony komórkowej, dzięki czemu
zachowuje ona selektywną przepuszczalność (m.in. nie wypuszczając
enzymów na zewnątrz, a jonów wapnia do komórki).
Dystrofina, która zawiera mutacje jest niestabilna, co powoduje zaburzenia w
utrzymywaniu prawidłowych interakcji pomiędzy dystrofiną a aktyną,
przyczyniając się jednocześnie do zaburzeń w strukturze całego cytoszkieletu
i powodując zmiany w budowie błony biologicznej.
Dochodzi do przechodzenia enzymów (kinaza kreatynowa, adolaza,
transaminazy) na zewnątrz błony komórkowej. Powoduje to wzrost ich
poziomu, w surowicy, znacznie powyżej normy.
Do komórki wnikają natomiast bez przeszkód jony wapnia, co powoduje jej
martwicę, stymulację fibroblastów i rozrost tkanki włóknistej w miejscu tkanki
mięśniowej.
W ten sposób z biegiem czasu coraz więcej włókien jest traconych, prowadzi
do degeneracji tkanki mięśniowej.
Dystrofina składa się z 3684 aminokwasów i ma masę 427 kDa.
Dystrofina jest białkiem kodowanym przez gen leżący na chromosomie X.
Rola dystrofiny polega na "uszczelnianiu" błony komórkowej, dzięki czemu
zachowuje ona selektywną przepuszczalność (m.in. nie wypuszczając
enzymów na zewnątrz, a jonów wapnia do komórki).
Dystrofina, która zawiera mutacje jest niestabilna, co powoduje zaburzenia w
utrzymywaniu prawidłowych interakcji pomiędzy dystrofiną a aktyną,
przyczyniając się jednocześnie do zaburzeń w strukturze całego cytoszkieletu
i powodując zmiany w budowie błony biologicznej.
Dochodzi do przechodzenia enzymów (kinaza kreatynowa, adolaza,
transaminazy) na zewnątrz błony komórkowej. Powoduje to wzrost ich
poziomu, w surowicy, znacznie powyżej normy.
Do komórki wnikają natomiast bez przeszkód jony wapnia, co powoduje jej
martwicę, stymulację fibroblastów i rozrost tkanki włóknistej w miejscu tkanki
mięśniowej.
W ten sposób z biegiem czasu coraz więcej włókien jest traconych, prowadzi
do degeneracji tkanki mięśniowej.

Bibliografia
Biochemia, Jeremy M. Berg, John L. Tymoczko,
Lubert Stryer, wydawnictwo naukowe PWN
Warszawa 2005
Biochemia Harpera, Franciszek Kokot,
Aleksander Koj, Andrzej Kozik, Tadeusz
Wilczok, Wydawnictwo lekarski PZWL
warszawa 2008
Fizjologia człowieka t.I, Stanisław Konturek,
Tomasz Brzozowski, Kraków 2003
Histologia , Wojciech Sawicki, Wydawnictwo
Lekarski PZWL, Warszawa 2003
Document Outline
- Slide 1
- Budowa miofibryli - sarkomer
- Miozyna
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Filamenty cienkie
- Aktyna
- Tropomiozyna
- Slide 12
- Troponina
- Slide 14
- Troponina I
- Troponina T
- Troponina C
- Skurcz mięśnia
- Kalmodulina
- Kaldesmon(87 kDa)
- Inne białka regulujące skurcz mięsni
- Kalcyneuryna
- Tytyna
- Nebulina
- Desmina
- Dystrfofina
- Bibliografia
Wyszukiwarka
Podobne podstrony:
Tk mięśniowa, białka kurczliwe, mech skurczu antastic pl
Białka uczestniczące w skurczu mięśni tomek
Analiza chodu mięśnie biorące udział w chodzie
1 Rodzaje pracy fizjologia skurczu mięśnia
Energetyka skurczu miesnia i wy Nieznany
Energetyka skurczu mięśnia plan prezentacji
Kości tworzą układ dźwigni poruszających się w następstwie skurczów mięśni szkieletowych
Regulacja siły skurczu mięśnia
bioenergetyka skurczu mięśnia
Potencjał spoczynkowy i czynnościowy nerwu, rejestracja skurczów mięśni szkieletowych po pobudzeniu
Mechanizm skurczu mięśnia szkieletowego
SZCZEGÓŁOWE ZASADY BEZPIECZEŃSTWA OSÓB BIORĄCYCH UDZIAŁ W IMPREZACH ŻEGLARSKICH, Edukacja w Sporcie,
rodzaje skurczów mięśniowych, Anatomia i fizjologia, anatomia
skurcz miesni, Biologia maturalna
7 Leki zwiększające siłę skurczu mięśnia sercowego1, 4farmacja, III koło
FIZJOLOGIA MIĘŚNIA SERCOWEGO – AKTYWNOŚĆ SKURCZOWA MIĘŚNIA SERCOWEGO - instrukcja, Medycyna i Zdrowi
więcej podobnych podstron