
FIZJOLOGIA MIĘŚNI
FIZJOLOGIA MIĘŚNI
LIANA PUCHALSKA, STANISŁAW
KOWALEWSKI
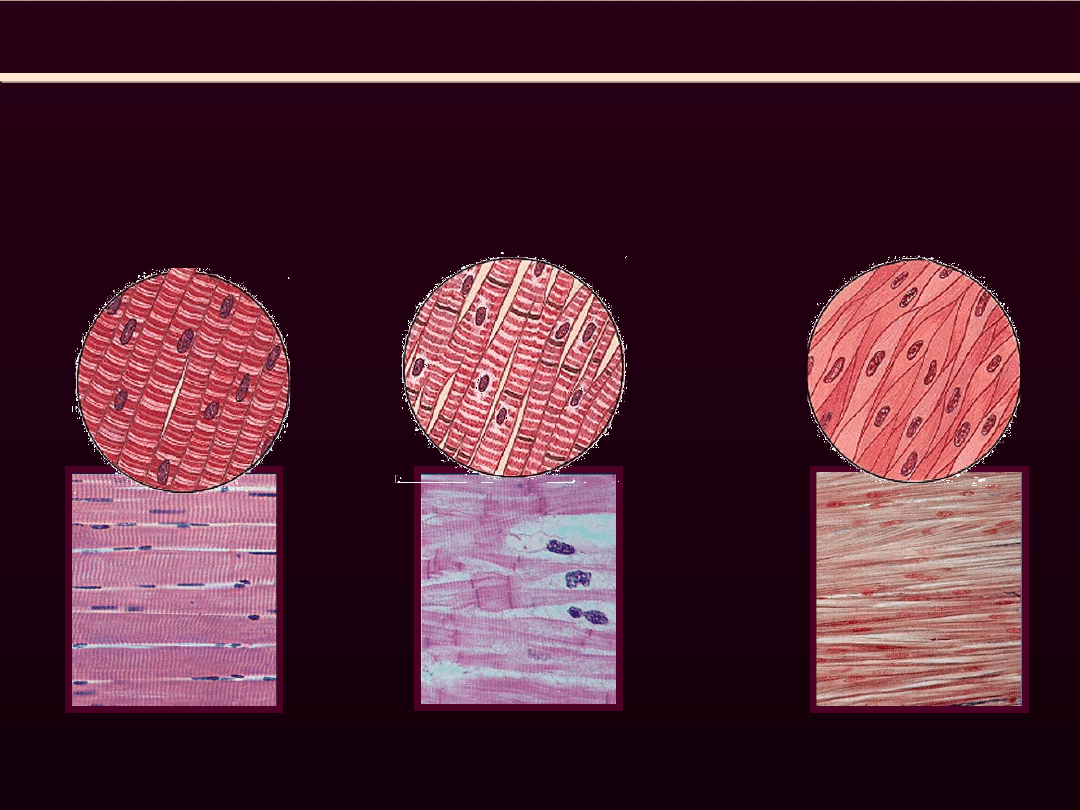
RODZAJE MIĘSNI
RODZAJE MIĘSNI
poprzecznie prążkowane
poprzecznie prążkowane
gładkie
gładkie
Mięsień szkieletowy
Mięsień szkieletowy
Mięsień sercowy
Mięsień sercowy
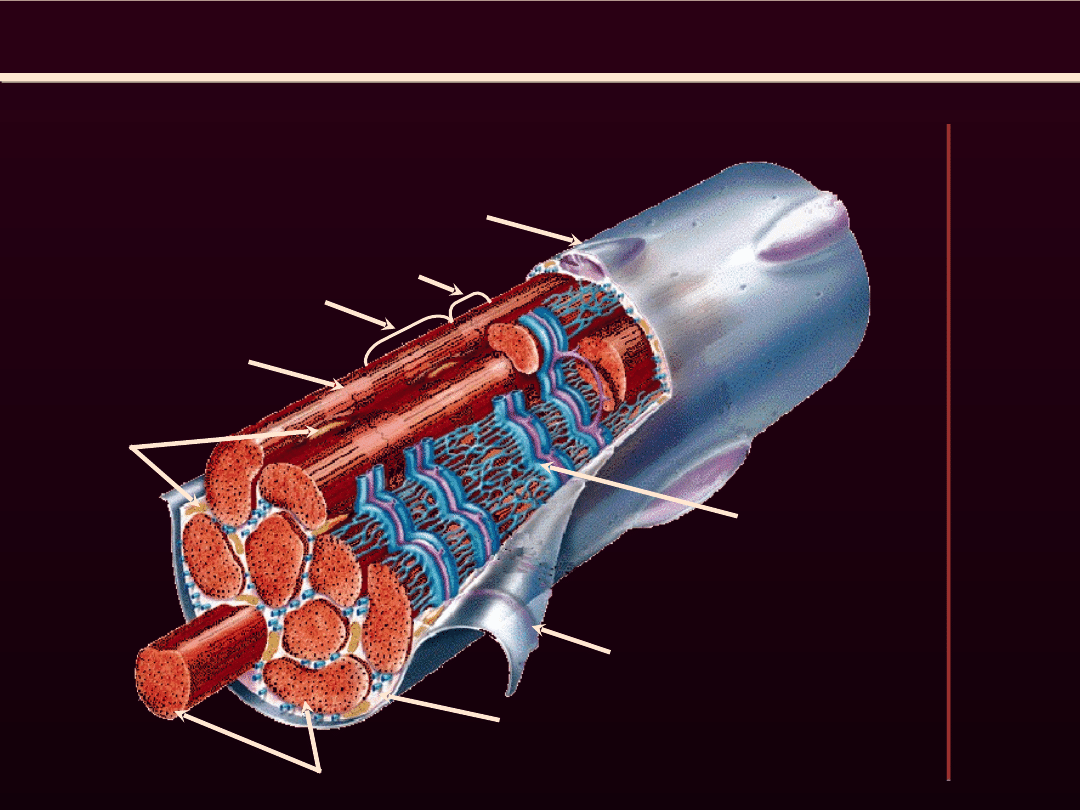
STRUKTURA WŁÓKNA
MIĘSNIOWEGO
STRUKTURA WŁÓKNA
MIĘSNIOWEGO
jądro
jądro
mitochondria
mitochondria
błona graniczna Z
błona graniczna Z
prążek I
prążek I
prążek A
prążek A
miofibryle
miofibryle
sarkoplazma
sarkoplazma
sarkolema
sarkolema
Siateczka śród-
plazmatyczna
Siateczka śród-
plazmatyczna

MIĘSNIE SZKIELETOWE
. Budowa sarkomeru
MIĘSNIE SZKIELETOWE
. Budowa sarkomeru
IZOTROPOWY
IZOTROPOWY
ANIZOTROPOWY
H
M
aktyna
F
miozyn
a
Z
Z
Z
Z
Aktyna F jest polimerem aktyny G. Każdy
monomer ma miejsce wiążące główki miozyny.
W stanie spoczynku miejsca te są zasłonięte
przez kompleks tropomiozynowo-troponinowy
Aktyna F jest polimerem aktyny G. Każdy
monomer ma miejsce wiążące główki miozyny.
W stanie spoczynku miejsca te są zasłonięte
przez kompleks tropomiozynowo-troponinowy
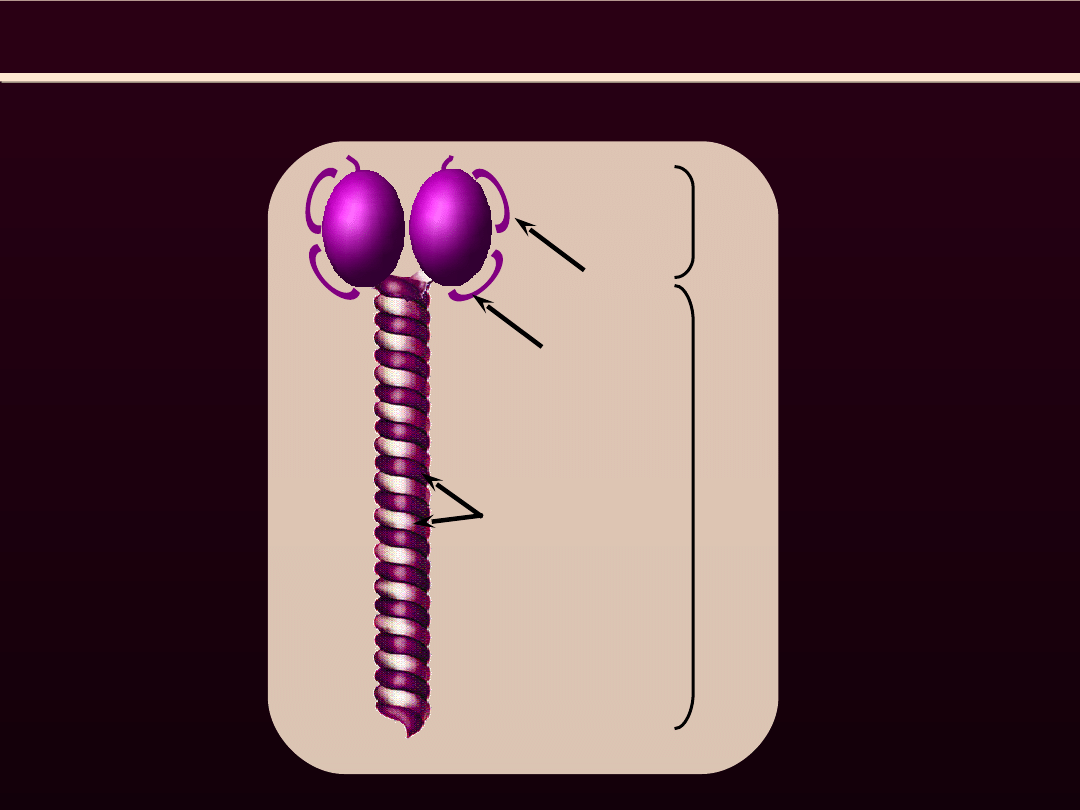
MIOZYNA
MIOZYNA
G
G
C
C
G
G
N
N
N
N
łańcuch ciężki H
łańcuch ciężki H
łańcuch lekki
podstawowy L
p
łańcuch lekki
podstawowy L
p
łańcuch lekki
regulatorowy L
r
łańcuch lekki
regulatorowy L
r
T
R
Z
O
N
T
R
Z
O
N
G
Ł
Ó
W
K
A
G
Ł
Ó
W
K
A
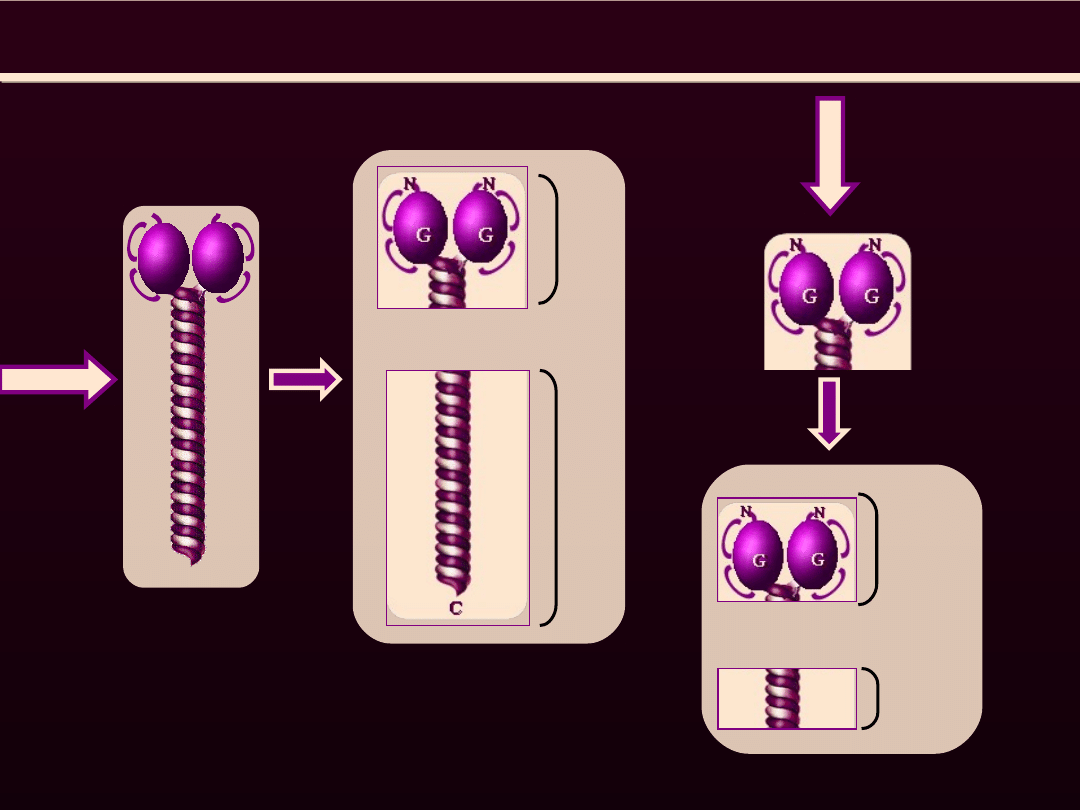
TRAWIENIE
MIOZYNY
TRAWIENIE
MIOZYNY
G
G
C
C
G
G
N
N
N
N
TRYPSYNA
m
e
ro
m
io
zy
n
a
c
ię
żk
a
(
H
M
M
)
m
e
ro
m
io
zy
n
a
le
k
k
a
(
L
M
M
)
P
A
P
A
IN
A
H
M
M
S
-1
a
k
ty
w
n
o
ść
A
T
P
-
a
zy
H
M
M
S
-2

AKTYNA
AKTYNA
- podjednostka T wiąże się z tropomiozyną
- podjednostka T wiąże się z tropomiozyną
- aktyna G
- aktyna G
- tropomiozyna
- tropomiozyna
- troponina
- troponina
- podjednostka I hamuje aktywność ATP-azową aktomiozyny
- podjednostka I hamuje aktywność ATP-azową aktomiozyny
- podjednostka C wiąże jony Ca
2+
i znosi hamujące
działanie podjednostki I
- podjednostka C wiąże jony Ca
2+
i znosi hamujące
działanie podjednostki I

BIAŁKA WSPOMAGAJACE KOMÓRKI MIĘŚNIOWEJ
BIAŁKA WSPOMAGAJACE KOMÓRKI MIĘŚNIOWEJ
Białka
pozasarkomerowej
części
cytoszkieletu
DESMINA
–
utrzymuje
poprzeczne
uporządkowanie
w
miofibrilach
DYSTROFINA
– łączy α-aktyninę błony granicznej Z z
przezbłono-wym kompleksem glikoproteinowym, który z kolei
tworzy połącze-nia z elementami sprężystymi macierzy
pozakomórkowej
Białka podporowe sarkomeru
α-AKTYNINA
– wchodzi w skład błony granicznej Z i umozliwia
zakotwiczenia do niej aktyny, konektyny (tininy) i nebuliny
KONEKTYNA
– koniec N zakotwiczony jest w błonie granicznej
Z, a koniec C w prążku M. fragment w obrębie prążka I ma
kształt sprężysty, a w obrębie prążka A ma powtarzające się
domeny wiążące LMM
NEBULINA –
owinięta wokół nitek aktyny utrzymując ich stałą
długość. Tego białka nie ma w mięśniu sercowym
Białka
pozasarkomerowej
części
cytoszkieletu
DESMINA
–
utrzymuje
poprzeczne
uporządkowanie
w
miofibrilach
DYSTROFINA
– łączy α-aktyninę błony granicznej Z z
przezbłono-wym kompleksem glikoproteinowym, który z kolei
tworzy połącze-nia z elementami sprężystymi macierzy
pozakomórkowej
Białka podporowe sarkomeru
α-AKTYNINA
– wchodzi w skład błony granicznej Z i umozliwia
zakotwiczenia do niej aktyny, konektyny (tininy) i nebuliny
KONEKTYNA
– koniec N zakotwiczony jest w błonie granicznej
Z, a koniec C w prążku M. fragment w obrębie prążka I ma
kształt sprężysty, a w obrębie prążka A ma powtarzające się
domeny wiążące LMM
NEBULINA –
owinięta wokół nitek aktyny utrzymując ich stałą
długość. Tego białka nie ma w mięśniu sercowym
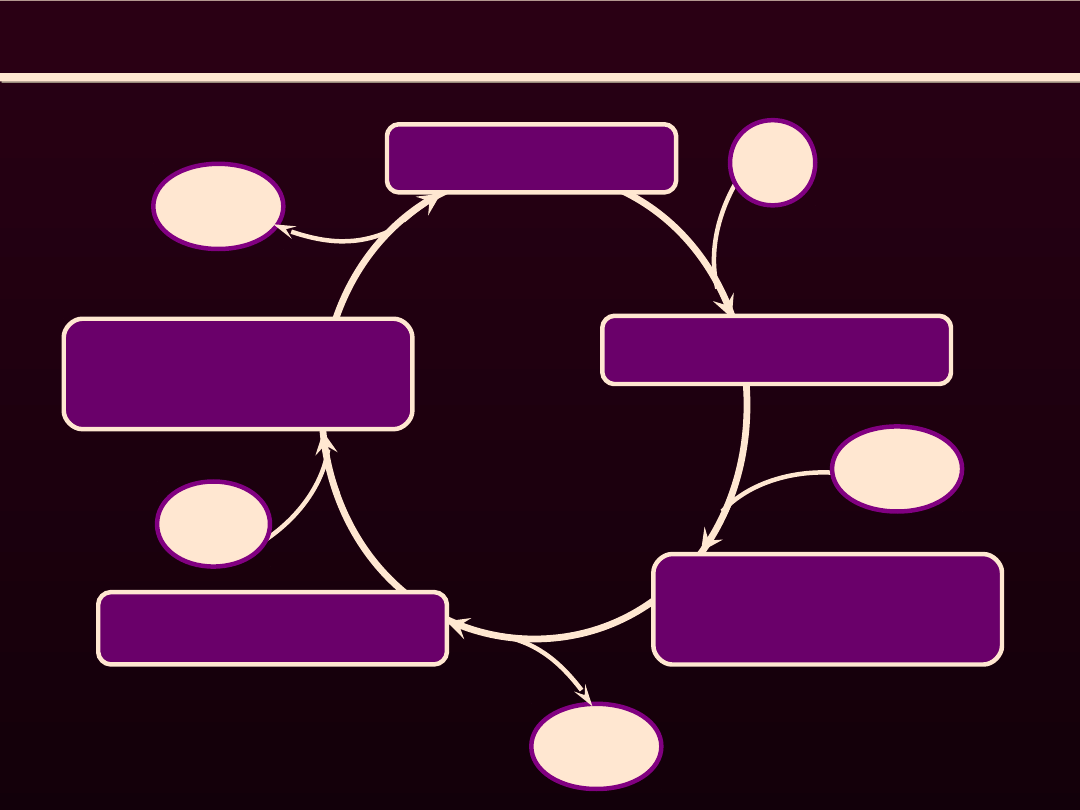
CYKL MOSTKA
CYKL MOSTKA
ATP-miozyna
ATP-miozyna
ADP-P
i
-
miozyna
ADP-P
i
-
miozyna
H
2
O
H
2
O
aktyna
aktyna
aktyna-miozyna
ADP-P
i
aktyna-miozyna
ADP-P
i
aktyna-miozyna
aktyna-miozyna
ADP+P
i
ADP+P
i
aktyna-miozyna
ATP
aktyna-miozyna
ATP
ATP
ATP
aktyna
aktyna
CYKL MOSTKA
CYKL MOSTKA
Pobudzenie sarkolemy
Pobudzenie sarkolemy
Wzrost stężenia Ca
2+
w sarkoplazmie
Wzrost stężenia Ca
2+
w sarkoplazmie
Obrót polimerów tropomiozynowych o 25° wokół filamentów
aktynowych
Obrót polimerów tropomiozynowych o 25° wokół filamentów
aktynowych
Częściowe odsłonięcie miejsc wiązania główek miozyny i częściowe
słabe wiązanie miozyny z aktyną
Częściowe odsłonięcie miejsc wiązania główek miozyny i częściowe
słabe wiązanie miozyny z aktyną
Wzrost powinowactwa podjednostki C do Ca
2+
Wzrost powinowactwa podjednostki C do Ca
2+
Dalszy obrót polimerów tropomiozynowych wokół filamentów
aktynowych
Dalszy obrót polimerów tropomiozynowych wokół filamentów
aktynowych
Całkowite odsłonięcie miejsc wiązania główek miozyny i silne
wiązanie miozyny z aktyną
Całkowite odsłonięcie miejsc wiązania główek miozyny i silne
wiązanie miozyny z aktyną
Wraz z odłączeniem P
i
dochodzi do zmiany kąta miedzy główką a
resztą mostka i przesunięcia aktyny względem miozyny
Wraz z odłączeniem P
i
dochodzi do zmiany kąta miedzy główką a
resztą mostka i przesunięcia aktyny względem miozyny
Wymiana ADP na ATP kończy cykl mostka
Wymiana ADP na ATP kończy cykl mostka
A
A

CYKL MOSTKA
CYKL MOSTKA
Podczas skurczu mięśnia zachodzi
wiele cykli mostków
Im
większe
jest
obciążenie
mięśnia,
tym
bardziej
zsynchronizowane muszą być cykle
mostków
w
obrębie
jednego
sarkomeru, jak i w sarko-merach
sąsiadujących
Podczas skurczu mięśnia zachodzi
wiele cykli mostków
Im
większe
jest
obciążenie
mięśnia,
tym
bardziej
zsynchronizowane muszą być cykle
mostków
w
obrębie
jednego
sarkomeru, jak i w sarko-merach
sąsiadujących

SPRĘŻENIE ELEKTRO-MECHANICZNE
SPRĘŻENIE ELEKTRO-MECHANICZNE
Przekazywanie
pobudzenia
wywołującego
powstanie
potencjału
czynnościowego w komórce mięśniowej
i prowadzącego do skracania się
sarkomerów nazywane jest sprężeniem
elektromechanicznym
Potencjał spoczynkowy mięśnia szkiele-
towego wynosi od -85 do -90 mV. Za tak
niski potencjał odpowiedzialny jest
niespecyficzny kanał kationowy, który
jest 100 razy bardziej przepuszczalny
dla jonów K
+
niż dla jonów Na
+
Przekazywanie
pobudzenia
wywołującego
powstanie
potencjału
czynnościowego w komórce mięśniowej
i prowadzącego do skracania się
sarkomerów nazywane jest sprężeniem
elektromechanicznym
Potencjał spoczynkowy mięśnia szkiele-
towego wynosi od -85 do -90 mV. Za tak
niski potencjał odpowiedzialny jest
niespecyficzny kanał kationowy, który
jest 100 razy bardziej przepuszczalny
dla jonów K
+
niż dla jonów Na
+

Uwolnienie acetylocholiny (ACh) w płytce motorycznej
Uwolnienie acetylocholiny (ACh) w płytce motorycznej
Otwarcie kanałów ACh zależnych i przesunięcie wartości potencjału
sarkolemy do -65 mV
Otwarcie kanałów ACh zależnych i przesunięcie wartości potencjału
sarkolemy do -65 mV
Otwarcie kanałów Na
+
-potencjałozaleznych
Otwarcie kanałów Na
+
-potencjałozaleznych
Wzrost potencjału błonowego do -40 mV pod wpływem prądu
sodowego
Wzrost potencjału błonowego do -40 mV pod wpływem prądu
sodowego
Otwarcie kanałów Ca
2+
typu L w błonie kanalików poprzecznych
(kanał receptor dihydropirydynowy)
Otwarcie kanałów Ca
2+
typu L w błonie kanalików poprzecznych
(kanał receptor dihydropirydynowy)
Wzrost stężenia Ca
2+
w sarkoplazmie
Wzrost stężenia Ca
2+
w sarkoplazmie
Zmiana konfiguracji białka JFP
Zmiana konfiguracji białka JFP
Uwolnienie Ca
2+
z siateczki sarkoplazmatycznej przez kanał-receptor
rianodynowy
Uwolnienie Ca
2+
z siateczki sarkoplazmatycznej przez kanał-receptor
rianodynowy
Znaczny wzrost stężenia Ca
2+
w sarkoplazmie
Znaczny wzrost stężenia Ca
2+
w sarkoplazmie
SPRĘŻENIE ELEKTRO-MECHANICZNE
SPRĘŻENIE ELEKTRO-MECHANICZNE
Cykl mostka
Cykl mostka
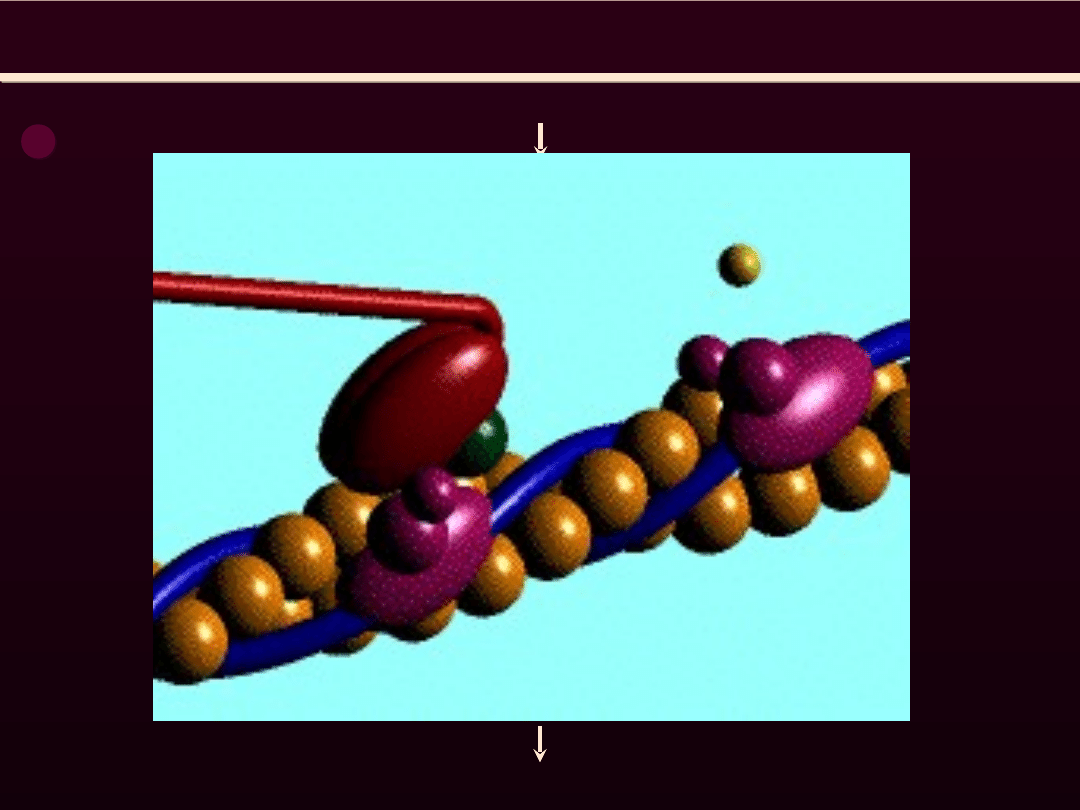
SIATECZKA SARKOPLAZMATYCZNA
SIATECZKA SARKOPLAZMATYCZNA
Część zbiornikowa
SS
Część cewkowa SS
zawiera Ca
2+
ATP-azę
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+ L
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
-
kalsekwestrina
Ca
2+
-
kalsekwestrina
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Stopkę łączącą stanowią:
- kanał Ca
2+
zbiornika końcowego SS (receptor rianodynowy)
- 4 kanały Ca
2+
(receptor dihydropirydynowy)
- białko JFP ( białko wiążące stopki)
Stopkę łączącą stanowią:
- kanał Ca
2+
zbiornika końcowego SS (receptor rianodynowy)
- 4 kanały Ca
2+
(receptor dihydropirydynowy)
- białko JFP ( białko wiążące stopki)
Stopka łącząca
JF
P
JF
P
JF
P
Ca
2+
-
kalsekwestrina
Ca
2+
-
kalsekwestrina
Ca
2+
-
kalsekwestrina
Ca
2+
-
kalsekwestrina

ROZKURCZ MIĘŚNIA
ROZKURCZ MIĘŚNIA
Rozkurcz mięśnia następuje po za-
kończeniu cyklu mostków
Warunkiem rozkurczu jest zmniej-
szenie
stężenia
Ca
2+
w
sarkoplazmie poniżej 10
-7
mol/L.
Dochodzi do tego na skutek
uruchamiania
(zniesienia
hamowania) pompy wapniowej
Ca
2+
-ATPazy oraz usuwanie Ca
2+
przez wymiennik 3Na
+
/Ca
2+
Rozkurcz mięśnia następuje po za-
kończeniu cyklu mostków
Warunkiem rozkurczu jest zmniej-
szenie
stężenia
Ca
2+
w
sarkoplazmie poniżej 10
-7
mol/L.
Dochodzi do tego na skutek
uruchamiania
(zniesienia
hamowania) pompy wapniowej
Ca
2+
-ATPazy oraz usuwanie Ca
2+
przez wymiennik 3Na
+
/Ca
2+

SKURCZ MIĘŚNIA SZKIELETOWEGO
SKURCZ MIĘŚNIA SZKIELETOWEGO
Skurcz pojedynczy
Skurcz pojedynczy
Skurcz tężcowy
Skurcz tężcowy
zupełny
zupełny
niezupełn
y
niezupełn
y
Wyróżniamy
Wyróżniamy
Czas trwania potencjału czynnościowego w
mięśniu szkieletowym wynosi od 5 ms do 10
ms
Czas trwania skurczu w mięśniu szkie-
letowym wynosi od 7,5 ms do 120 ms
Czas trwania potencjału czynnościowego w
mięśniu szkieletowym wynosi od 5 ms do 10
ms
Czas trwania skurczu w mięśniu szkie-
letowym wynosi od 7,5 ms do 120 ms
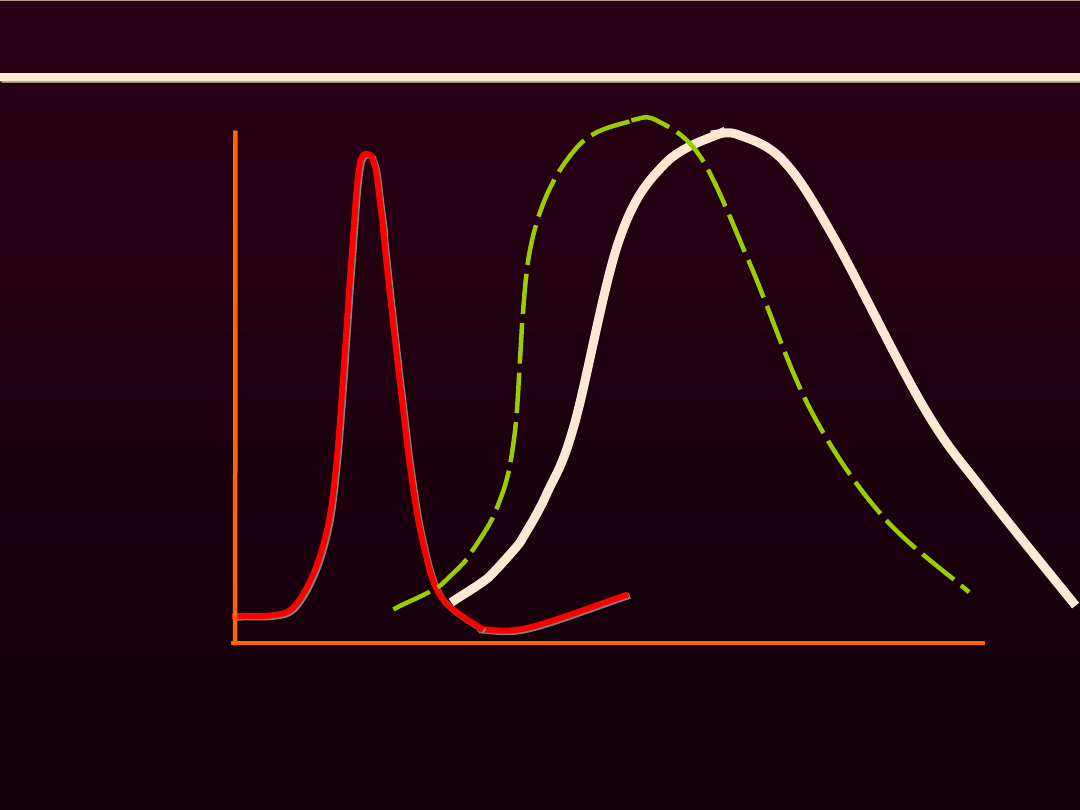
SKURCZ MIĘŚNIA SZKIELETOWEGO
POJEDYNCZY
SKURCZ MIĘŚNIA SZKIELETOWEGO
POJEDYNCZY
5
5
10
10
0
0
100%
100%
50%
50%
0%
0%
ms
ms
15
15
20
20
Depolaryzacja i refrakcja wygasają w czasie powstawania
wstępującego ramienia skurczu pojedynczego
Depolaryzacja i refrakcja wygasają w czasie powstawania
wstępującego ramienia skurczu pojedynczego
potencjał
czynnościowy
potencjał
czynnościowy
Skurcz mięśnia
Skurcz mięśnia
[Ca
2+
]
[Ca
2+
]
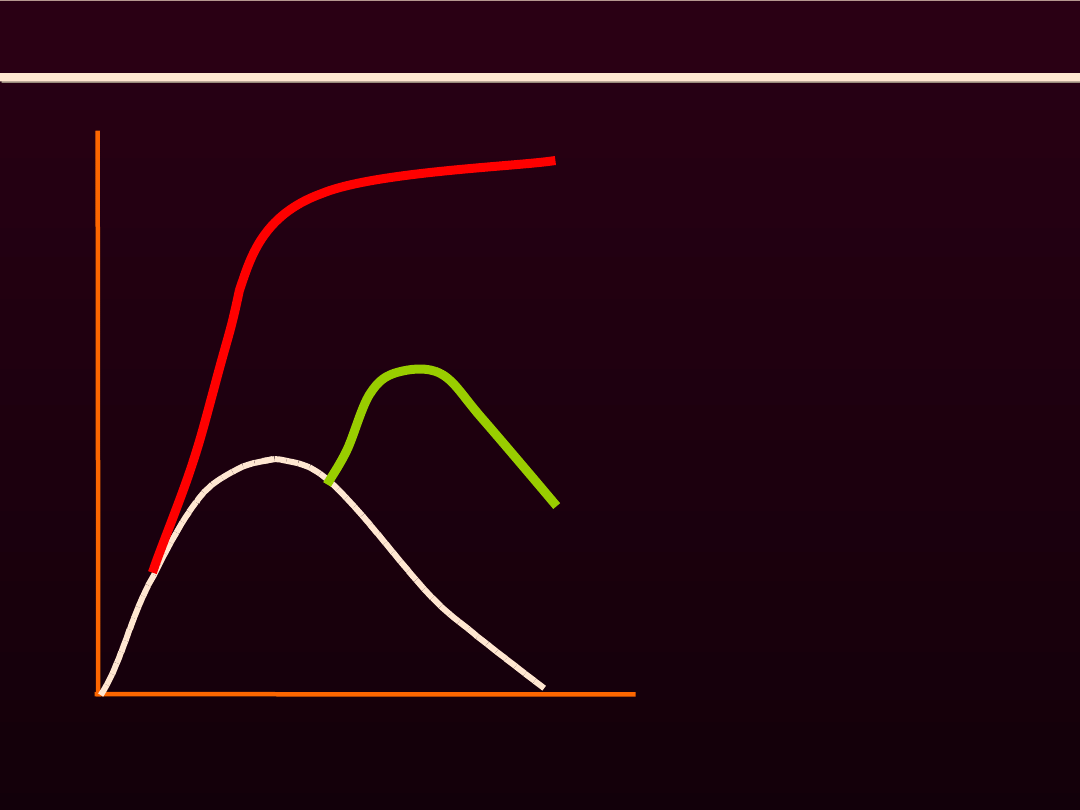
SKURCZ MIĘŚNIA SZKIELETOWEGO TĘŻCOWY
SKURCZ MIĘŚNIA SZKIELETOWEGO TĘŻCOWY
S
IŁ
A
S
K
U
R
C
Z
U
S
IŁ
A
S
K
U
R
C
Z
U
CZAS
TRWANIA
SKURCZU
CZAS
TRWANIA
SKURCZU
skurcz
pojedynczy
skurcz
pojedynczy
skurcz
tężcowy
niezupełny
skurcz
tężcowy
niezupełny
skurcz
tężcowy
zupełny
skurcz
tężcowy
zupełny
jeżeli każde następne
pobudzenie występuje
wtedy, gdy mięsień za-
czyna się rozkurczać,
mó-wimy
o
skurczu
tężco-wym niezupełnym
Jeżeli każde kolejne
pobudzenie przypada na
ramie
wstępujące,
mówi-my
o
skurczu
tężcowym zupełnym
jeżeli każde następne
pobudzenie występuje
wtedy, gdy mięsień za-
czyna się rozkurczać,
mó-wimy
o
skurczu
tężco-wym niezupełnym
Jeżeli każde kolejne
pobudzenie przypada na
ramie
wstępujące,
mówi-my
o
skurczu
tężcowym zupełnym
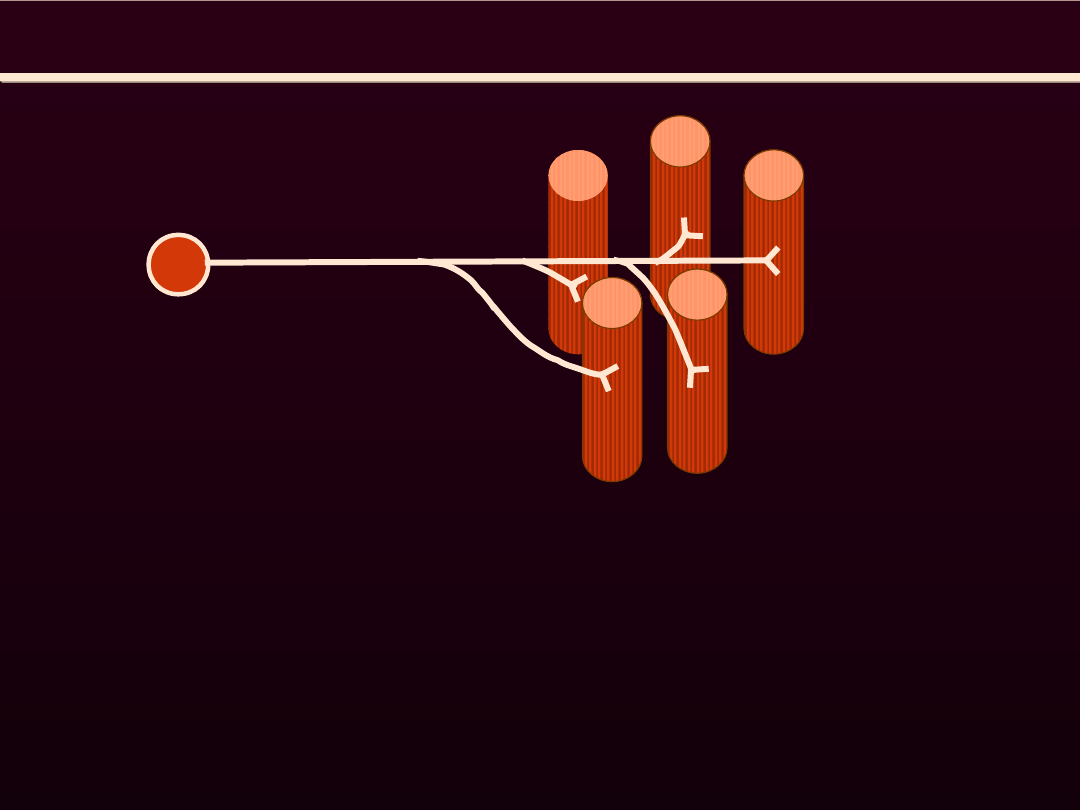
JEDNOSTKA MOTORYCZNA
JEDNOSTKA MOTORYCZNA
Jednostką motoryczna nazywamy motoneuron i
wszystkie komórki mięśniowe przez niego
unerwiane
Wszystkie komórki mięśniowe wchodzące w
skład tej samej jednostki motorycznej są tego
samego typu metabolicznego
Jednostką motoryczna nazywamy motoneuron i
wszystkie komórki mięśniowe przez niego
unerwiane
Wszystkie komórki mięśniowe wchodzące w
skład tej samej jednostki motorycznej są tego
samego typu metabolicznego

PODZIAŁ
JEDNOSTEK MOTORYCZNYCH
PODZIAŁ
JEDNOSTEK MOTORYCZNYCH
I
• małe motoneurony
• wysoka pobudliwość moto-
neuronów
• szybkie przewodzenie we
włóknach nerwowych
• mała liczba komórek mięś-
niowych
•
głównie
komórki
mięśniowe czerwone
I
• małe motoneurony
• wysoka pobudliwość moto-
neuronów
• szybkie przewodzenie we
włóknach nerwowych
• mała liczba komórek mięś-
niowych
•
głównie
komórki
mięśniowe czerwone
II
• duże motoneurony
• niska pobudliwość moto-
neuronów
•
bardzo
szybkie
przewodzenie we włóknach
nerwowych
• dużą liczba komórek mięś-
niowych
•
głównie
komórki
mięśniowe białe
II
• duże motoneurony
• niska pobudliwość moto-
neuronów
•
bardzo
szybkie
przewodzenie we włóknach
nerwowych
• dużą liczba komórek mięś-
niowych
•
głównie
komórki
mięśniowe białe

REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO
REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO
Prawo „wszystko albo nic”
Prawo „wszystko albo nic”
Dotyczy pojedynczych włókien szybkich
białych mięśni szkieletowych. Skurcz poje-
dynczych włókien mięśniowych następuje
tylko
po
zadziałaniu
bodźca
nadprogowego
Siła
skurczu
białego
mięśnia
szkieletowego zależy od intensywności
bodźca. Odpowiedź „wszystko albo nic”
występuje tylko w miejscach położonych
blisko miejsca działania bodźca. Wzrost
intensywności
bodźca
prowadzi
do
równomiernego pobudzenia wszystkich
włókien mięśnia
Dotyczy pojedynczych włókien szybkich
białych mięśni szkieletowych. Skurcz poje-
dynczych włókien mięśniowych następuje
tylko
po
zadziałaniu
bodźca
nadprogowego
Siła
skurczu
białego
mięśnia
szkieletowego zależy od intensywności
bodźca. Odpowiedź „wszystko albo nic”
występuje tylko w miejscach położonych
blisko miejsca działania bodźca. Wzrost
intensywności
bodźca
prowadzi
do
równomiernego pobudzenia wszystkich
włókien mięśnia

REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO
REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO
Prawo „wszystko albo nic” nie oznacza, że
odpowiedź pobudzanego włókna mięśniowego
będzie zawsze jednakowa. Siła skurczu mięśnia w
skurczu tężco-wym niezupełnym będzie większa,
niż w skurczu pojedynczym, natomiast w skurczu
tężcowym zupełnym siła skurczu mięśnia jest
jeszcze
większa.
Siła
skurczu
zależy
od
częstotliwości pobudzenia
Jeśli mięsień znajduje się w rozkurczu tuż po
skurczu
tężcowym
zupełnym,
pojedyncze
pobudzenie nadprogowe może wywołać skurcz
silniejszy od poprzedniego.
W warunkach niedotlenienia zmniejsza się nie
tylko siła skurczu, wydłużeniu ulega tez czas
trwania
rozkurczu.
Gdy
zasoby
ATP
są
wyczerpane
mięsień
wcale
nie
może
się
rozkurczyć
Prawo „wszystko albo nic” nie oznacza, że
odpowiedź pobudzanego włókna mięśniowego
będzie zawsze jednakowa. Siła skurczu mięśnia w
skurczu tężco-wym niezupełnym będzie większa,
niż w skurczu pojedynczym, natomiast w skurczu
tężcowym zupełnym siła skurczu mięśnia jest
jeszcze
większa.
Siła
skurczu
zależy
od
częstotliwości pobudzenia
Jeśli mięsień znajduje się w rozkurczu tuż po
skurczu
tężcowym
zupełnym,
pojedyncze
pobudzenie nadprogowe może wywołać skurcz
silniejszy od poprzedniego.
W warunkach niedotlenienia zmniejsza się nie
tylko siła skurczu, wydłużeniu ulega tez czas
trwania
rozkurczu.
Gdy
zasoby
ATP
są
wyczerpane
mięsień
wcale
nie
może
się
rozkurczyć

SKURCZ MIĘŚNIA SZKIELETOWEGO
SKURCZ MIĘŚNIA SZKIELETOWEGO
Podział
skurczów
względem
zmiany
długości mięśnia i generowania siły
Podział
skurczów
względem
zmiany
długości mięśnia i generowania siły
Skurcz izotoniczny
Występuje
wtedy,
gdy
mięsień może się skraca
ale nie generuje napięcia,
bo nie są rozciągane
elementy sprężyste
Skurcz izotoniczny
Występuje
wtedy,
gdy
mięsień może się skraca
ale nie generuje napięcia,
bo nie są rozciągane
elementy sprężyste
Skurcz izometryczny
Występuje
wtedy,
gdy
mięsień nie może się
skracać.
Generuje
napięcie,
ponieważ
sarkomery skra-cają się
kosztem
rozcią-gania
elementów
sprężys-tych
ułożonych szeregowo.
Skurcz izometryczny
Występuje
wtedy,
gdy
mięsień nie może się
skracać.
Generuje
napięcie,
ponieważ
sarkomery skra-cają się
kosztem
rozcią-gania
elementów
sprężys-tych
ułożonych szeregowo.
Czysty skurcz izotoniczny nie występuje nigdy, a czysty
skurcz izometryczny rzadko
Najczęściej występują skurcze dwufazowe - auksotoniczne
Czysty skurcz izotoniczny nie występuje nigdy, a czysty
skurcz izometryczny rzadko
Najczęściej występują skurcze dwufazowe - auksotoniczne
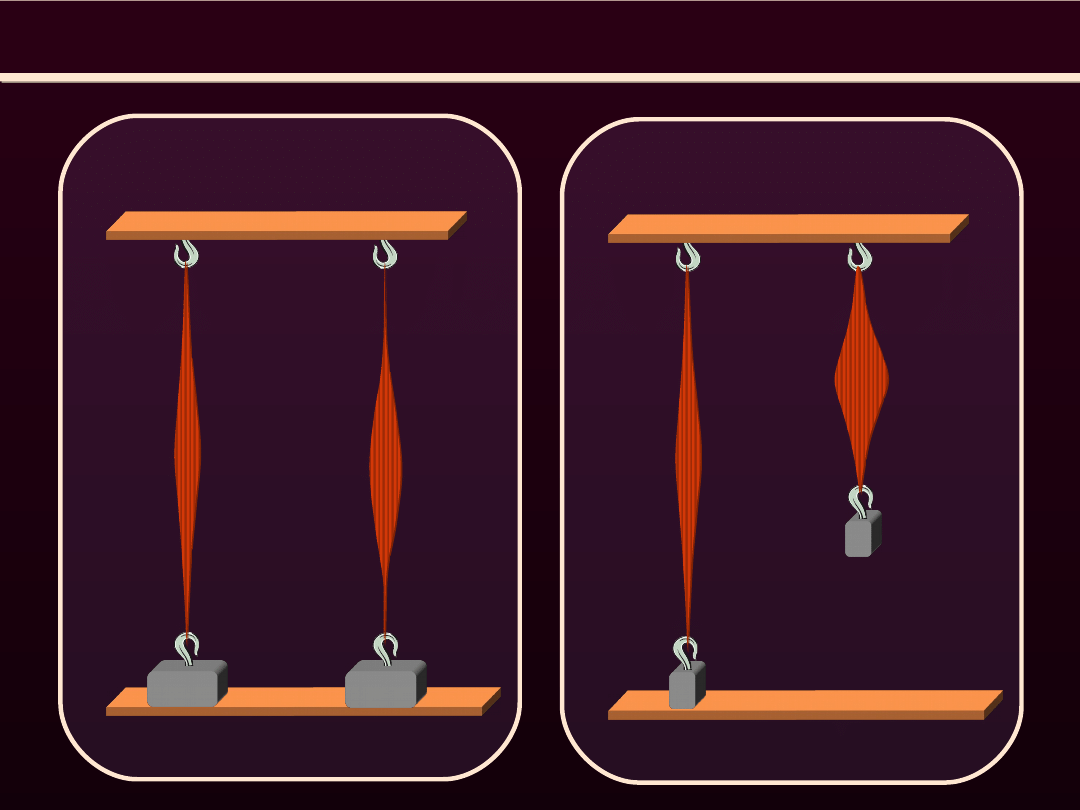
SKURCZ MIĘŚNIA SZKIELETOWEGO
SKURCZ MIĘŚNIA SZKIELETOWEGO
spoczynek skurcz izotoniczny
spoczynek skurcz izotoniczny
Skurcz izotoniczny
Skurcz izotoniczny
spoczynek skurcz izometryczny
spoczynek skurcz izometryczny
Skurcz izometryczny
Skurcz izometryczny
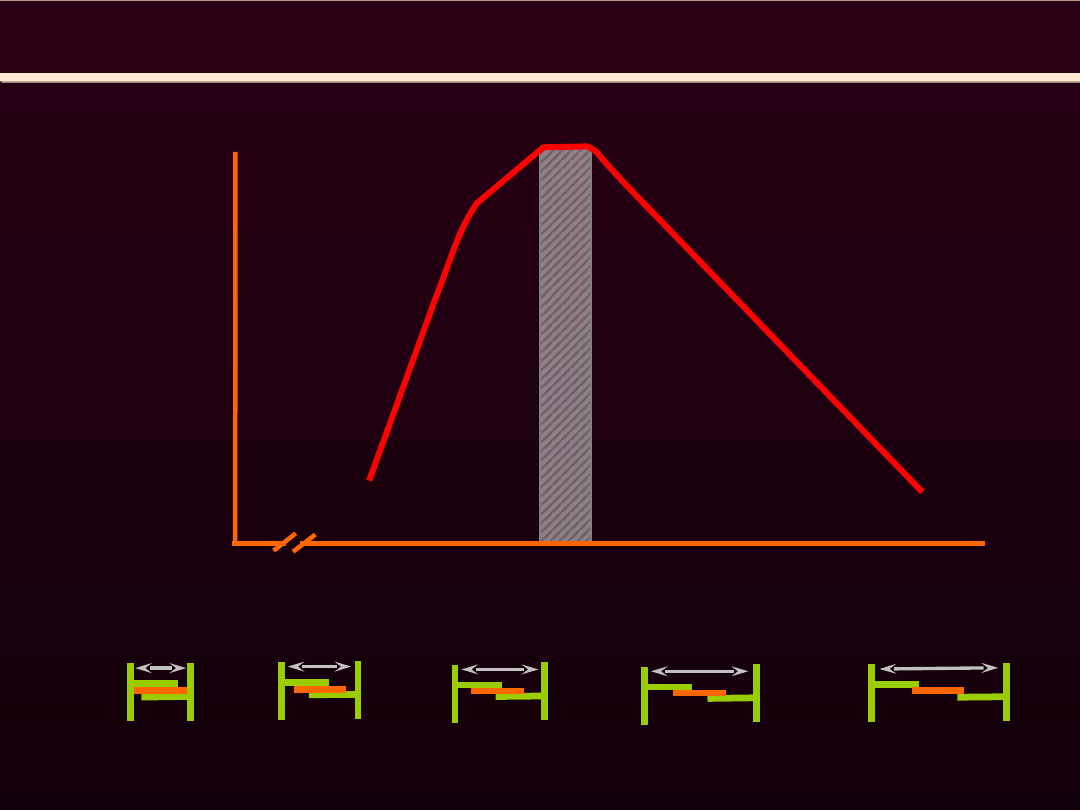
REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO
REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO
1.2 1.4 1.6 1.8 2.0 2.2 2.4 2.6 2.8 3.0 3.2 3.4 3.6
1.2 1.4 1.6 1.8 2.0 2.2 2.4 2.6 2.8 3.0 3.2 3.4 3.6
100%
100%
40%
40%
0%
0%
długość sarkomeru (μm)
długość sarkomeru (μm)
80%
80%
20%
20%
60%
60%
N
a
p
ię
c
ie
i
zo
m
e
tr
y
c
zn
e
(%
m
a
k
s
im
u
m
)
N
a
p
ię
c
ie
i
zo
m
e
tr
y
c
zn
e
(%
m
a
k
s
im
u
m
)
Z
a
k
re
s
m
a
k
s
y
m
a
ln
e
g
o
n
a
p
ię
c
ia
Z
a
k
re
s
m
a
k
s
y
m
a
ln
e
g
o
n
a
p
ię
c
ia
1.65
1.65
1.90
1.90
2.05
2.05
2.20
2.20
3.65
3.65
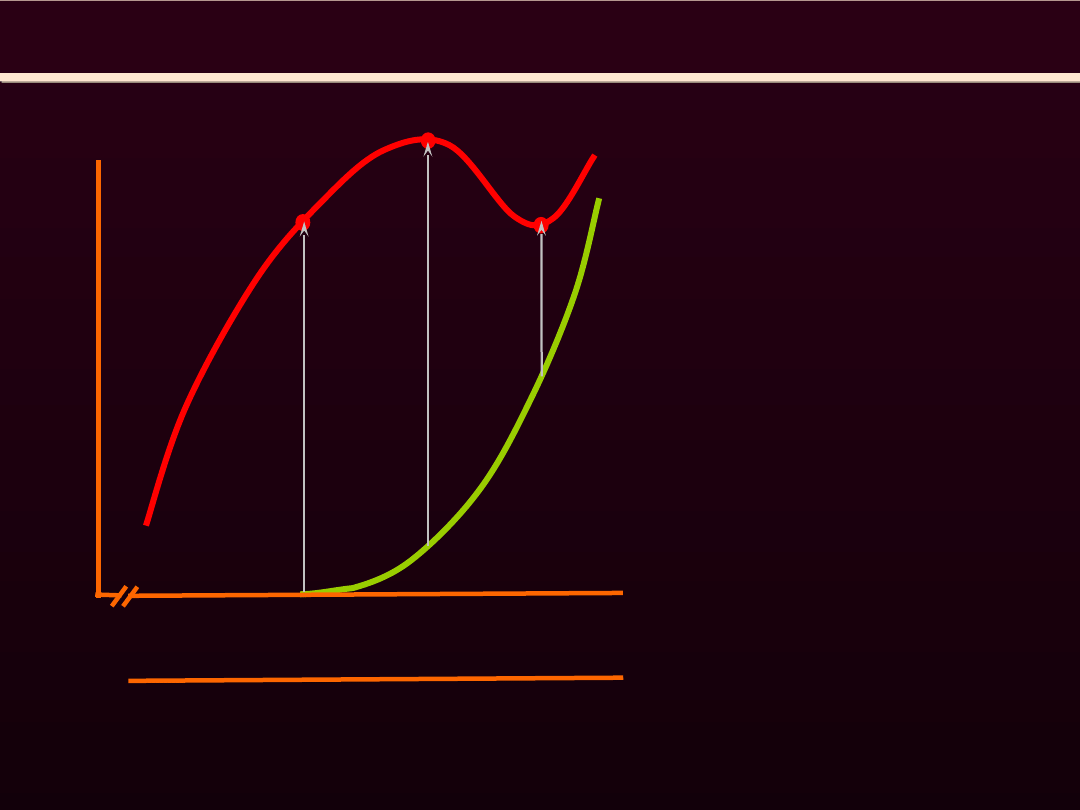
REGULACJA SIŁY
SKURCZU MIĘŚNIA SZKIELETOWEGO
REGULACJA SIŁY
SKURCZU MIĘŚNIA SZKIELETOWEGO
80 90 100
80 90 100
100%
100%
40%
40%
0%
0%
względna długość mięśnia
(100% = długość przy sile maksymalnej)
względna długość mięśnia
(100% = długość przy sile maksymalnej)
80%
80%
20%
20%
60%
60%
S
ił
a
m
ię
ś
n
ia
(%
o
d
m
a
k
s
im
u
m
w
s
k
u
rc
z
u
i
z
o
m
e
tr
y
c
z
n
y
m
)
S
ił
a
m
ię
ś
n
ia
(%
o
d
m
a
k
s
im
u
m
w
s
k
u
rc
z
u
i
z
o
m
e
tr
y
c
z
n
y
m
)
napięcie całkowite
napięcie całkowite
napięcie bierne
napięcie bierne
1.8 2.0 2.2
1.8 2.0 2.2
długość sarkomeru (μm)
długość sarkomeru (μm)
n
a
p
ię
c
ie
c
zy
n
n
e
n
a
p
ię
c
ie
c
zy
n
n
e
Wypadkowa napięcia
całkowitego jest sumą
napięcia
biernego
i
czynnego
Sprężystość mięśnia
zale-ży
od
elementów
sprężys-tych położonych
szeregowo
(elementy
tkanki łącznej, odcinki
szyjkowe
miozyny)
i
równolegle (błona włókna
mięśniowego,
tkanka
łącz-na) oraz elementów
kurczli-wych. Rozciąganie
tych
ele-mentów
jest
przyczyną
na-pięcia
biernego
Napięcie czynne jest
określone
przez
ilość
most-ków
aktynowo-
miozyno-wych i dlatego
zmienia
się
wraz
z
długością mięśnia
Wypadkowa napięcia
całkowitego jest sumą
napięcia
biernego
i
czynnego
Sprężystość mięśnia
zale-ży
od
elementów
sprężys-tych położonych
szeregowo
(elementy
tkanki łącznej, odcinki
szyjkowe
miozyny)
i
równolegle (błona włókna
mięśniowego,
tkanka
łącz-na) oraz elementów
kurczli-wych. Rozciąganie
tych
ele-mentów
jest
przyczyną
na-pięcia
biernego
Napięcie czynne jest
określone
przez
ilość
most-ków
aktynowo-
miozyno-wych i dlatego
zmienia
się
wraz
z
długością mięśnia

REGULACJA SZYBKOŚCI SKRACANIA MIĘŚNIA SZKIELETOWEGO
REGULACJA SZYBKOŚCI SKRACANIA MIĘŚNIA SZKIELETOWEGO
Maksymalna (bez obciążenia) szybkość skracania
sarkomeru jest równa maksymalnej prędkości
ślizgania aktyny i miozyny względem siebie. Tym
szybciej odbywa się to przesuwanie im większa jest
aktywność ATP-azowa miozyny
Ponieważ sarkomery w miofibrylach położone są
szeregowo, skracania ich są sumowane, więc przy
takiej samej szybkości skracania sarkomeru długi
mięsień będzie skracał się szybciej, niż mięsień
krótki
Szybkość skracania obciążonego mięśnia w
skurczu izotonicznym będzie zależała od stopnia
obciążenia
Maksymalna (bez obciążenia) szybkość skracania
sarkomeru jest równa maksymalnej prędkości
ślizgania aktyny i miozyny względem siebie. Tym
szybciej odbywa się to przesuwanie im większa jest
aktywność ATP-azowa miozyny
Ponieważ sarkomery w miofibrylach położone są
szeregowo, skracania ich są sumowane, więc przy
takiej samej szybkości skracania sarkomeru długi
mięsień będzie skracał się szybciej, niż mięsień
krótki
Szybkość skracania obciążonego mięśnia w
skurczu izotonicznym będzie zależała od stopnia
obciążenia
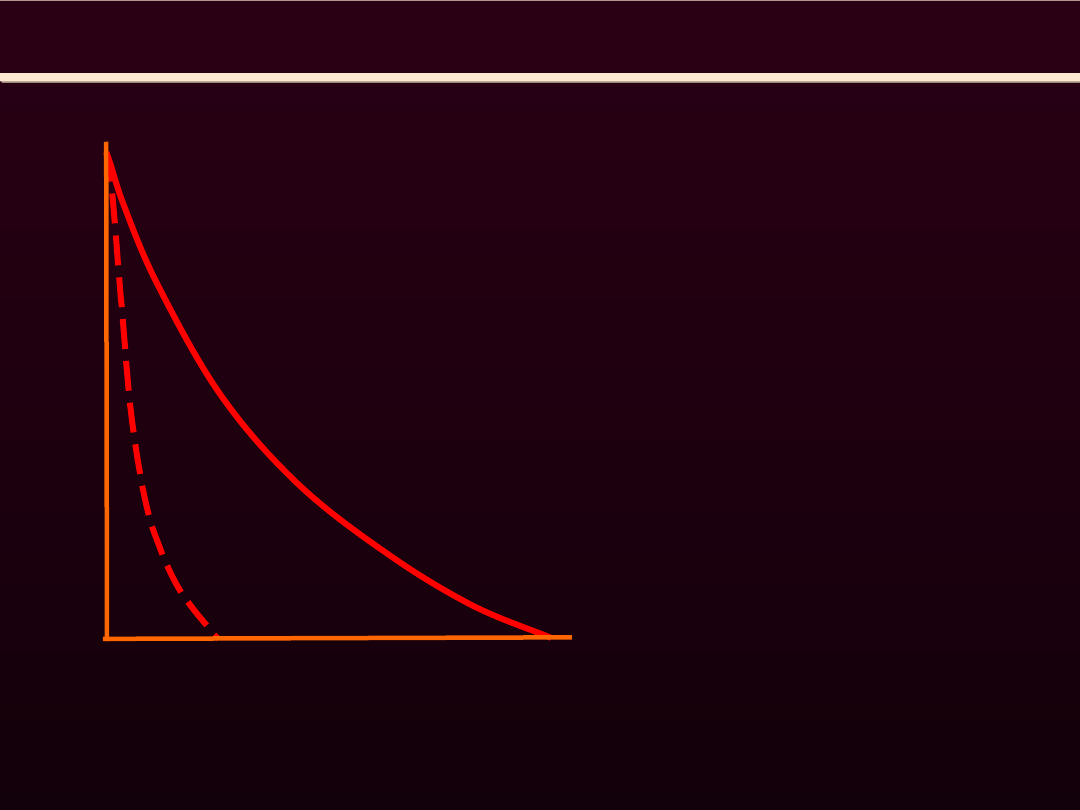
REGULACJA SZYBKOŚCI SKRACANIA MIĘŚNIA SZKIELETOWEGO
REGULACJA SZYBKOŚCI SKRACANIA MIĘŚNIA SZKIELETOWEGO
0
25
50
75
100
0
25
50
75
100
S
zy
b
k
o
ść
sk
ra
c
a
n
ia
(%
V
m
a
x
)
S
zy
b
k
o
ść
sk
ra
c
a
n
ia
(%
V
m
a
x
)
Siła mięśniowa (mN)
Siła mięśniowa (mN)
100
75
50
25
0
100
75
50
25
0
długie włókna
mięśniowe
krótkie włókna
mięśniowe
Szybkość skracania mięśnia
zmniejsza
się
wraz
ze
zwiększeniem obciążenia. Gdy
obciążenie jest równe połowie
siły maksymalnej osiąganej w
warunkach
izometrycznych,
szybkość skracania osiąga ok.
1/5
wartości
maksymalnie
możliwej (obserwowanej bez
obciążenia)
Szybko
skracający
się
mięsień rozwija mniejszą siłę
niż mięsień skracający się
wolniej
lub
poprzednio
rozciągnięty. Duży ciężar jeśli
w ogóle może być podniesiony,
to tylko wolno. Szybkie ruchy
natomiast możliwe tylko wtedy,
gdy nie wymagają dużej siły

METABOLIZM ENERGETYCZNY MIĘŚNI SZKIELETOWYCH
METABOLIZM ENERGETYCZNY MIĘŚNI SZKIELETOWYCH
Zapas ATP (5 μmol/g mięśnia) w komórce
mięśniowej jest wystarczający na 1 do 2 sekund
skurczu
∆G ≈ 50 kJ (12 kcal)/mol ATP
Substratami energetycznymi, wykorzystywanymi
przez mięśnie do resyntezy ATP, są:
- fosfokreatyna
- glikogen mięśniowy
- glukoza wychwytywana z krwi
- WKT
- ketokwasy
- aminokwasy
Zapas ATP (5 μmol/g mięśnia) w komórce
mięśniowej jest wystarczający na 1 do 2 sekund
skurczu
∆G ≈ 50 kJ (12 kcal)/mol ATP
Substratami energetycznymi, wykorzystywanymi
przez mięśnie do resyntezy ATP, są:
- fosfokreatyna
- glikogen mięśniowy
- glukoza wychwytywana z krwi
- WKT
- ketokwasy
- aminokwasy
METABOLIZM ENERGETYCZNY MIĘŚNI SZKIELETOWYCH
METABOLIZM ENERGETYCZNY MIĘŚNI SZKIELETOWYCH
•
Fosfokreatyna
-
zapas CrP ok. 25
μmol/g
mięśnia wystarcza na 50 skurczów. Energia CrP
umożliwia krótkotrwały (10 do 20s) intensywny
wysiłek)
kreatyna + ADP
kreatyna + ATP
•
Synteza ATP z 2 częsteczek ADP
ADP + ADP
ATP + AMP
•
Fosfokreatyna
-
zapas CrP ok. 25
μmol/g
mięśnia wystarcza na 50 skurczów. Energia CrP
umożliwia krótkotrwały (10 do 20s) intensywny
wysiłek)
kreatyna + ADP
kreatyna + ATP
•
Synteza ATP z 2 częsteczek ADP
ADP + ADP
ATP + AMP
P
i
kinaza
adenylanowa
kinaza
adenylanowa

METABOLIZM ENERGETYCZNY MIĘŚNI SZKIELETOWYCH
METABOLIZM ENERGETYCZNY MIĘŚNI SZKIELETOWYCH
•
Glikoliza beztlenowa
- zapas glukozy ok.
100 μmol/g mięśnia. Glikoliza
beztlenowa jest opóźniona w stosunku do rozpadu
CrP o ok. 30 s. Z 1 mola reszt glukozowych uzyskuje
się netto 2 mole ATP
Glikogen ATP +
kwas mlekowy
•
Glikoliza tlenowa
– jest znacznie bardziej
energetycznie wydajna. Z 1 mola glukozy na tej
drodze powstaje netto 38 moli ATP (czasem 36 moli
ATP)
Długotrwały wysiłek mięśniowy jest możliwy
tylko dzięki pozyskiwaniu energii z glukozy i
tłuszczów
•
Glikoliza beztlenowa
- zapas glukozy ok.
100 μmol/g mięśnia. Glikoliza
beztlenowa jest opóźniona w stosunku do rozpadu
CrP o ok. 30 s. Z 1 mola reszt glukozowych uzyskuje
się netto 2 mole ATP
Glikogen ATP +
kwas mlekowy
•
Glikoliza tlenowa
– jest znacznie bardziej
energetycznie wydajna. Z 1 mola glukozy na tej
drodze powstaje netto 38 moli ATP (czasem 36 moli
ATP)
Długotrwały wysiłek mięśniowy jest możliwy
tylko dzięki pozyskiwaniu energii z glukozy i
tłuszczów
glukozo-6-fosforan
glukozo-6-fosforan
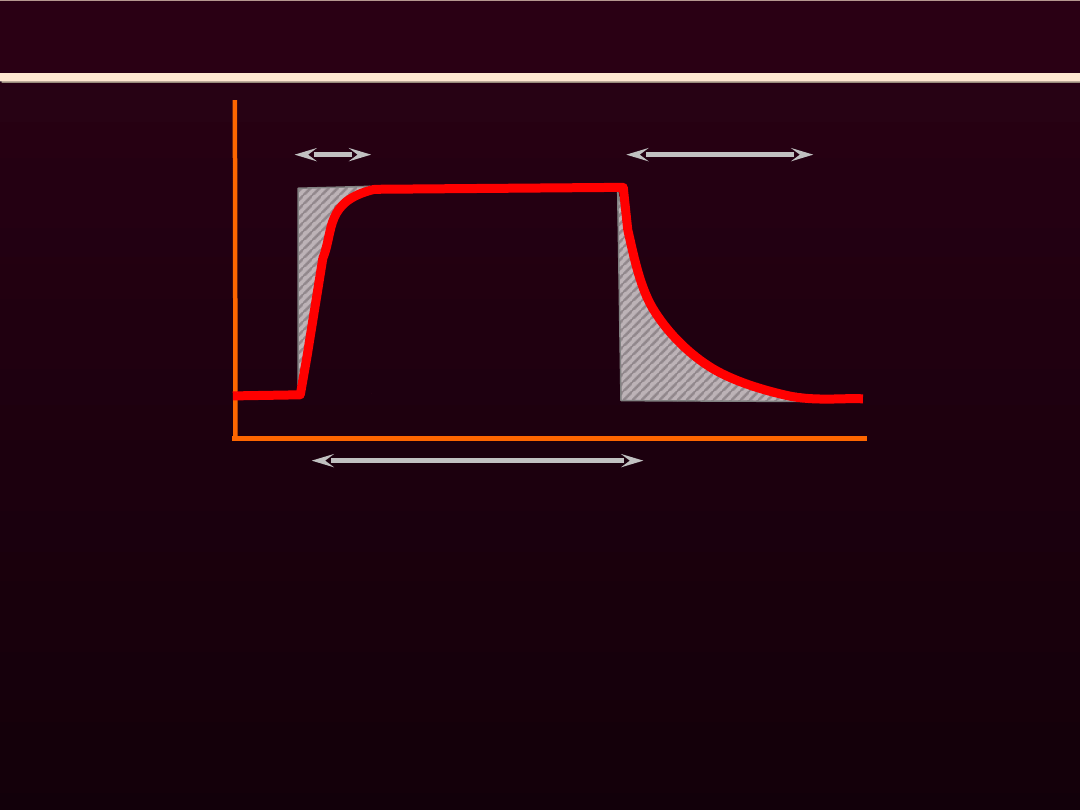
METABOLIZM ENERGETYCZNY MIĘŚNI SZKIELETOWYCH
METABOLIZM ENERGETYCZNY MIĘŚNI SZKIELETOWYCH
V
o
2
V
o
2
WYSIŁEK
WYSIŁEK
•
•
Deficyt
tlenowy
Deficyt
tlenowy
Dług
tlenowy
Dług
tlenowy
Zanim nastąpi aktywacja przemian tlenowych w mitochondriach
a czynność układu krążenia i oddechowego osiągnie poziom
odpowiadający zapotrzebowaniu tlenowemu, skurcz mięsni odbywa
się dzięki energii pochodzącej z rozpadu CrP i beztlenowej przemiany
glukozy. Jest to okres
DEFICYTU TLENOWEGO
, trwający zwykle kilka
minut (3-6 min).
Zaciągnięty
DŁUG TLENOWY
musi być oddany w okresie spoczynku
po wysiłku. A zatem zużycie tlenu, mimo spoczynku organizmu,
pozostaje jeszcze przez jakiś czas podwyższone
Zanim nastąpi aktywacja przemian tlenowych w mitochondriach
a czynność układu krążenia i oddechowego osiągnie poziom
odpowiadający zapotrzebowaniu tlenowemu, skurcz mięsni odbywa
się dzięki energii pochodzącej z rozpadu CrP i beztlenowej przemiany
glukozy. Jest to okres
DEFICYTU TLENOWEGO
, trwający zwykle kilka
minut (3-6 min).
Zaciągnięty
DŁUG TLENOWY
musi być oddany w okresie spoczynku
po wysiłku. A zatem zużycie tlenu, mimo spoczynku organizmu,
pozostaje jeszcze przez jakiś czas podwyższone

PODZIAŁ METABOLICZNY MIĘSNI
SZKIELETOWYCH
PODZIAŁ METABOLICZNY MIĘSNI
SZKIELETOWYCH
Typ I - czerwone
• przewaga metabolizmu
tle-nowego
•
duża
zawartość
mioglobiny
•
gęsta
sieć
naczyń
włosowa-tych
•
bardzo
dużo
mitochondriów
• powolne narastanie siły
skurczu
• powolne występowanie
zmę-czenia
Typ I - czerwone
• przewaga metabolizmu
tle-nowego
•
duża
zawartość
mioglobiny
•
gęsta
sieć
naczyń
włosowa-tych
•
bardzo
dużo
mitochondriów
• powolne narastanie siły
skurczu
• powolne występowanie
zmę-czenia
Typ II - białe
II a
• przewaga metabolizmu
bez-tlenowego
•
mała
zawartość
mioglobiny
•
mała
liczba
mitochondriów
• szybkie narastanie siły
skur-czu
•
szybkie
występowanie
zmę-czenia
II b
mają charakter pośredni
pomiędzy typem I i IIa
Typ II - białe
II a
• przewaga metabolizmu
bez-tlenowego
•
mała
zawartość
mioglobiny
•
mała
liczba
mitochondriów
• szybkie narastanie siły
skur-czu
•
szybkie
występowanie
zmę-czenia
II b
mają charakter pośredni
pomiędzy typem I i IIa

MIĘŚNIE GŁADKIE
MIĘŚNIE GŁADKIE
Skurcz mięśnia gładkiego nie podlega
świadomej kontroli
Mięśnie gładkie nie mają sarkomerów
Miozyna w mięśniu gładkim jest usytuowana na
całej długości aktyny
Skurcz
mięśnia
gładkiego
jest
wydajny
energetycznie ale powolny
Pomiędzy miocytami mięsni gładkich znajdują
się złącza szczelinowe
Wśród komórek mięsni gładkich są komórki
rozrusznikowe, cechujące się powstaniem w nich
samoistnej powolnej depolaryzacji
(w ścianie naczyń
krwionośnych znajdują się komórki mięśni gładkich o charakterze
rozrusznikowym, natomiast w ścianach żołądka i jelit znajdują się
komórki nerwowe Cajala)
Skurcz mięśnia gładkiego nie podlega
świadomej kontroli
Mięśnie gładkie nie mają sarkomerów
Miozyna w mięśniu gładkim jest usytuowana na
całej długości aktyny
Skurcz
mięśnia
gładkiego
jest
wydajny
energetycznie ale powolny
Pomiędzy miocytami mięsni gładkich znajdują
się złącza szczelinowe
Wśród komórek mięsni gładkich są komórki
rozrusznikowe, cechujące się powstaniem w nich
samoistnej powolnej depolaryzacji
(w ścianie naczyń
krwionośnych znajdują się komórki mięśni gładkich o charakterze
rozrusznikowym, natomiast w ścianach żołądka i jelit znajdują się
komórki nerwowe Cajala)

PORÓWNANIE MIĘŚNI GŁADKICH I MIĘSNI SZKIELETOWYCH
PORÓWNANIE MIĘŚNI GŁADKICH I MIĘSNI SZKIELETOWYCH
CECHY
CECHY
MIĘSIEŃ GŁADKI
MIĘSIEŃ GŁADKI
MIĘSIEŃ SZKIELETOWY
MIĘSIEŃ SZKIELETOWY
Włókna
Włókna
wrzecionowate
wrzecionowate
(maks
(maks
0.4
0.4
nm)
nm)
walcowate, długie
walcowate, długie
(maks
(maks
15
15
cm)
cm)
Mitochondri
Mitochondri
a
a
nieliczne
nieliczne
zależy od typu mięśnia
zależy od typu mięśnia
Jądro
Jądro
komórkowe
komórkowe
1
1
liczne
liczne
Sarkomery
Sarkomery
brak
brak
tak, maks. długość
tak, maks. długość
3.65
3.65
μ
μ
m
m
Syncytium
Syncytium
tak (złącza)
tak (złącza)
brak
brak
Siateczka
Siateczka
sarko-
sarko-
plazmatyczna
plazmatyczna
słabo rozwinięta
słabo rozwinięta
dobrze rozwinięta
dobrze rozwinięta
BUDOWA
BUDOWA

PORÓWNANIE MIĘŚNI GŁADKICH I MIĘSNI SZKIELETOWYCH
PORÓWNANIE MIĘŚNI GŁADKICH I MIĘSNI SZKIELETOWYCH
CECHY
CECHY
MIĘSIEŃ GŁADKI
MIĘSIEŃ GŁADKI
MIĘSIEŃ SZKIELETOWY
MIĘSIEŃ SZKIELETOWY
Charakter
Charakter
transmisji
transmisji
brak synapsy
brak synapsy
obecność synapsy
obecność synapsy
Troponina
Troponina
brak
brak
tak
tak
Regulacja cyklu
Regulacja cyklu
mostku
mostku
miozyna
miozyna
aktyna
aktyna
Stopień
Stopień
zależności od
zależności od
zewnątrz
zewnątrz
komórkowego
komórkowego
Ca
Ca
2+
2+
większy
większy
mniejszy
mniejszy
Kompleks
Kompleks
Ca
Ca
2+
2+
kalmodulin
kalmodulin
a
a
tak
tak
brak
brak
Komórki
Komórki
rozrusznikowe
rozrusznikowe
tak
tak
(czynność
(czynność
spontaniczna powolna)
spontaniczna powolna)
brak
brak
(konieczny jest bodziec
(konieczny jest bodziec
nerwowy)
nerwowy)
Skurcz tężcowy
Skurcz tężcowy
tak
tak
tak
tak
Zakres pracy
Zakres pracy
zmienna krzywa
zmienna krzywa
siła/długość
siła/długość
w maksimum krzywej
w maksimum krzywej
siły/długość
siły/długość
FUNKCJA
FUNKCJA
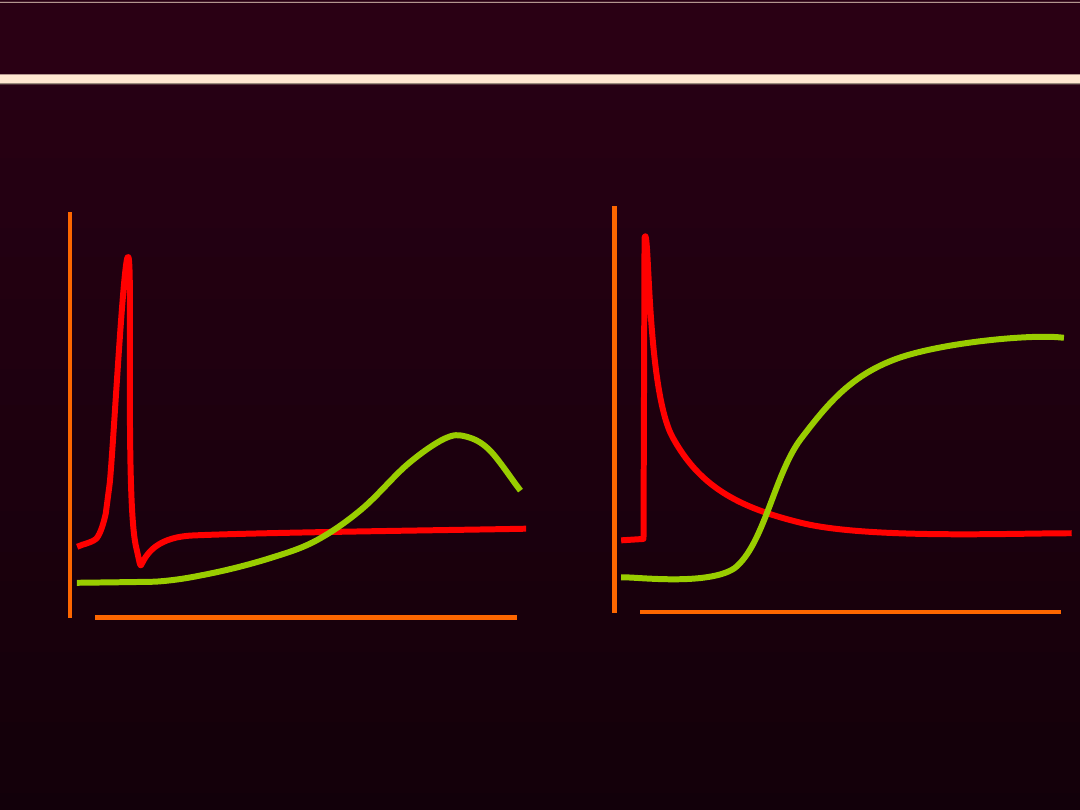
PORÓWNANIE MIĘŚNI GŁADKICH I MIĘSNI SZKIELETOWYCH
PORÓWNANIE MIĘŚNI GŁADKICH I MIĘSNI SZKIELETOWYCH
ODPOWIEDŹ NA BODZIEC
ODPOWIEDŹ NA BODZIEC
200
200
ms
ms
0
0
400
400
600
600
20
20
-60
-60
-20
-20
0
0
-40
-40
m
V
m
V
10
10
ms
ms
0
0
20
20
30
30
50
50
-100
-100
0
0
-50
-50
m
V
m
V
mięsień gładki
mięsień gładki
mięsień szkieletowy
mięsień szkieletowy

PODZIAŁ CZYNNOŚCIOWY MIĘŚNI GŁADKICH
PODZIAŁ CZYNNOŚCIOWY MIĘŚNI GŁADKICH
Wielojednostkowe
(mięśnie ściany nasieniowodów, rozwieracz źrenicy, mięśnie ścian zespoleń
tętniczo-żylnych w obrębie skóry)
obficie unerwione, mało wrażliwe na krążące we krwi
katecholaminy, nie mają własnego automatyzmu (nie ma
komórek rozrusznikowych)
Wielojednostkowe
(mięśnie ściany nasieniowodów, rozwieracz źrenicy, mięśnie ścian zespoleń
tętniczo-żylnych w obrębie skóry)
obficie unerwione, mało wrażliwe na krążące we krwi
katecholaminy, nie mają własnego automatyzmu (nie ma
komórek rozrusznikowych)
Jednostkowe
(mięśnie macicy i dużych naczyń tętniczych)
słabo unerwione, bardzo wrażliwe na krążące we krwi
katecholaminy, część z nich posiada własny automatyzm (są
wśród nich komórki rozrusznikowe)
Jednostkowe
(mięśnie macicy i dużych naczyń tętniczych)
słabo unerwione, bardzo wrażliwe na krążące we krwi
katecholaminy, część z nich posiada własny automatyzm (są
wśród nich komórki rozrusznikowe)
Typ pośredni
(mięśnie oskrzeli i pęcherza moczowego)
unerwione asymetrycznie, co wynika z różnej odległości pomiędzy
żylakowatościami zakończenia współczulnego a poszczególnymi
włóknami mięśnia
Typ pośredni
(mięśnie oskrzeli i pęcherza moczowego)
unerwione asymetrycznie, co wynika z różnej odległości pomiędzy
żylakowatościami zakończenia współczulnego a poszczególnymi
włóknami mięśnia

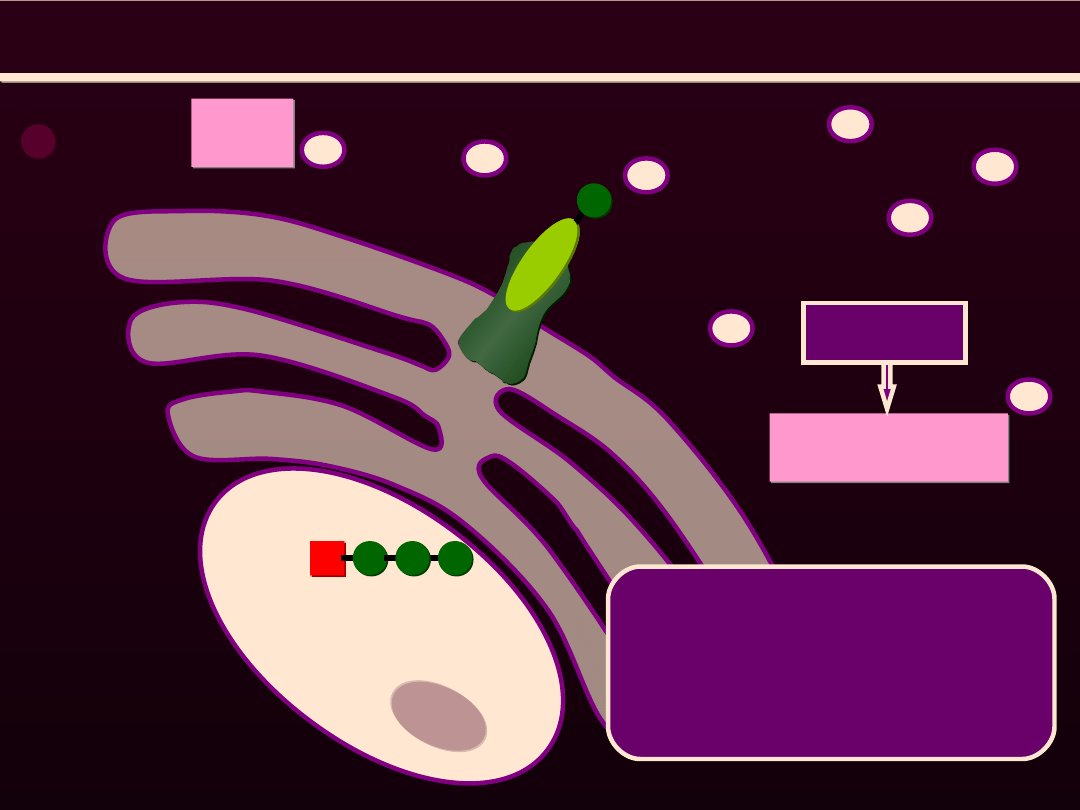
MECHANIZM SKURCZU MIĘŚNI GŁADKICH
MECHANIZM SKURCZU MIĘŚNI GŁADKICH
Wzrost stężenia Ca
2+
w sarkoplazmie
Wzrost stężenia Ca
2+
w sarkoplazmie
Uwolnienie Ca
2+
z siateczki sarkoplazmatycznej
Uwolnienie Ca
2+
z siateczki sarkoplazmatycznej
Jony Ca
2+
łączą się z kalmoduliną i aktywują zależną od
kompleksu Ca
2+
-kalmodulina kinazę, która fosforyluje L
p
główki miozyny
Jony Ca
2+
łączą się z kalmoduliną i aktywują zależną od
kompleksu Ca
2+
-kalmodulina kinazę, która fosforyluje L
p
główki miozyny
Znoszony jest wpływ hamujący L
p
na interakjcę aktyny i
miozyny
Znoszony jest wpływ hamujący L
p
na interakjcę aktyny i
miozyny
Cykl mostka (podobny do mięśnia szkieletowego)
Cykl mostka (podobny do mięśnia szkieletowego)
Bodziec nerwowy
lub
hormomalny
Bodziec nerwowy
lub
hormomalny
PKA
PKA
KINAZA ZALEŻNA
od C/Ca
2+
KINAZA ZALEŻNA
od C/Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
ATP-
aza
SERCA
fo
sf
ol
am
ba
n
fo
sf
ol
am
ba
n
KALMODULINA
KALMODULINA
Ca
2+
fo
sf
ol
am
ba
n
fo
sf
ol
am
ba
n
P
P
A
A
A
A
P
P
P
P
P
P
ATP-aza Ca
2+
części cewkowej SS
ATP-aza Ca
2+
części cewkowej SS
•
•
Bierze
udział
w
Bierze
udział
w
rozkurczu
mięśni
rozkurczu
mięśni
gładkich.
gładkich.
•
•
W mięśniu sercowym
W mięśniu sercowym
reguluje
tempo
i
reguluje
tempo
i
stopień rozkurczu
stopień rozkurczu
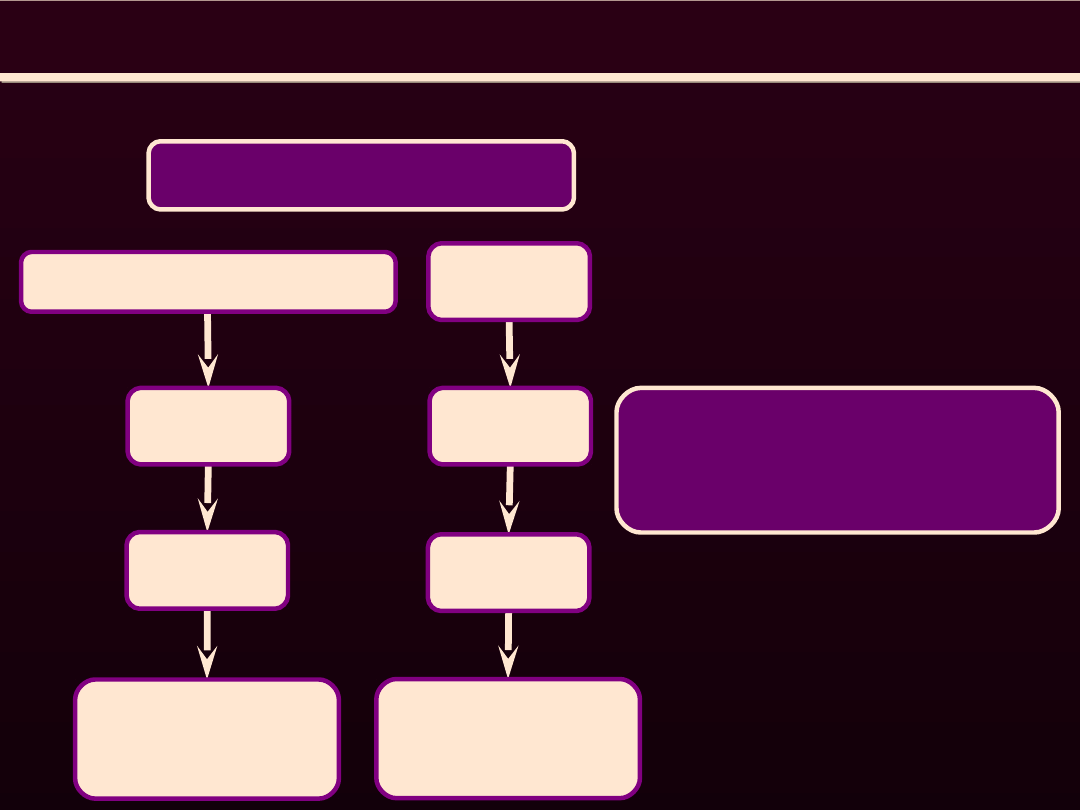
REGULACJA AKTYWNOŚCI
ATP-azy Ca
2+
REGULACJA AKTYWNOŚCI
ATP-azy Ca
2+
Wzrost aktywności
Wzrost aktywności
Hamują aktywność
wolne rodniki
tlenowe
Hamują aktywność
wolne rodniki
tlenowe
agonisci receptorów β
agonisci receptorów β
cAMP
cAMP
PKA
PKA
Fosforylacja
fosfolambanu
Fosforylacja
fosfolambanu
NO, ANP
NO, ANP
cGMP
cGMP
PKG
PKG
Fosforylacja
fosfolambanu
Fosforylacja
fosfolambanu
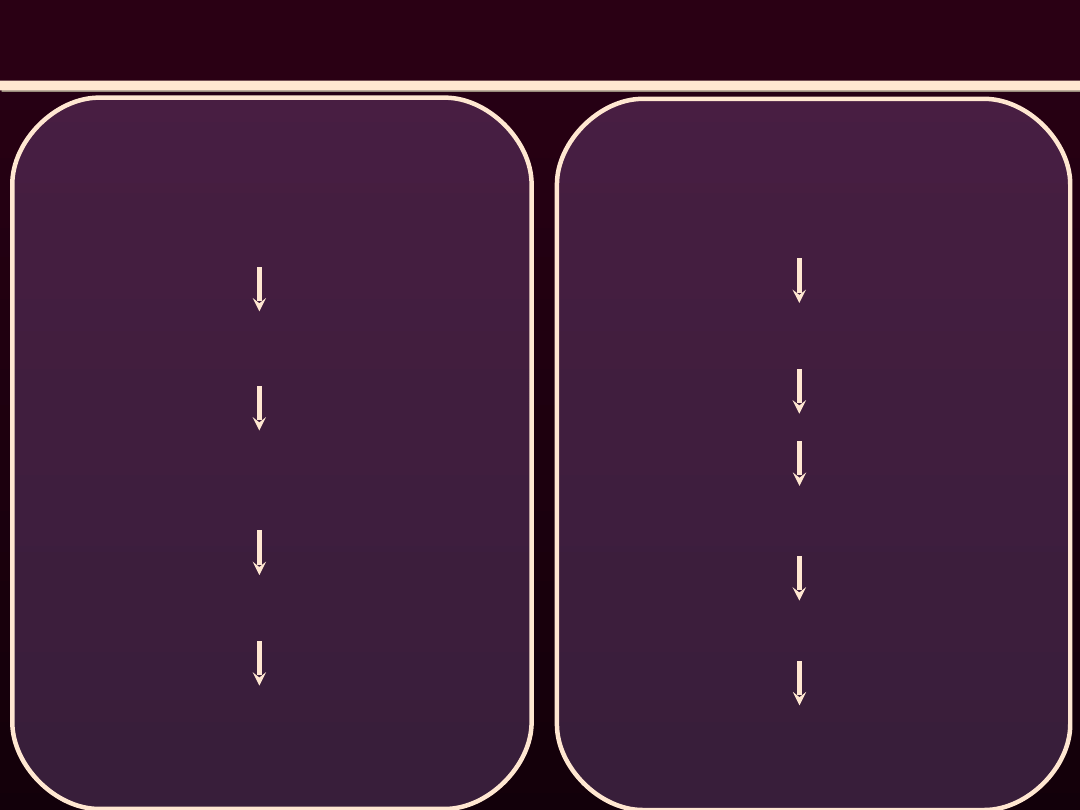
WPŁYW POBUDZENIA RECEPTORÓW ADRENERGICZNYCH NA
MIĘSNIE GŁADKIE NACZYŃ
WPŁYW POBUDZENIA RECEPTORÓW ADRENERGICZNYCH NA
MIĘSNIE GŁADKIE NACZYŃ
β-receptory
Pobudzenie receptora przez
noradrenalinę wydzialana z
zakończenia nerwowego
β-receptory
Pobudzenie receptora przez
noradrenalinę wydzialana z
zakończenia nerwowego
Wzrost stężenia cAMP w
sarkoplazmie
Wzrost stężenia cAMP w
sarkoplazmie
Aktywacja proteokinazy A (PKA)
Aktywacja proteokinazy A (PKA)
Fosforylacja kinazy zależnej od
kompleksu Ca2+-kalmodulina
Fosforylacja kinazy zależnej od
kompleksu Ca2+-kalmodulina
Zmniejszenie powinowactwa tej
kinazy
do kompleksu Ca
2+
-
kalmodulina
Zmniejszenie powinowactwa tej
kinazy
do kompleksu Ca
2+
-
kalmodulina
Zmniejszenie pobudliwości
skurczowej miocytu przy danym
stężeniu Ca
2+
Zmniejszenie pobudliwości
skurczowej miocytu przy danym
stężeniu Ca
2+
α
1
-receptory
Pobudzenie receptora przez
noradrenalinę wydzialana z
zakończenia nerwowego
α
1
-receptory
Pobudzenie receptora przez
noradrenalinę wydzialana z
zakończenia nerwowego
Wzrost stężenia DAG i IP
3
w
sarkoplazmie
Wzrost stężenia DAG i IP
3
w
sarkoplazmie
Wzrost szybkosci uwalniania Ca
2+
z SS oraz wzrost
przepuszczalnościn sarkolemy
dla jonów Ca
2+
Wzrost szybkosci uwalniania Ca
2+
z SS oraz wzrost
przepuszczalnościn sarkolemy
dla jonów Ca
2+
Przyspieszenie fosforylacji L
p
miozyny
Przyspieszenie fosforylacji L
p
miozyny
Skurcz mięsni gładkich naczyń
Skurcz mięsni gładkich naczyń
R
R
PKA
GDP
GDP
GDP
GDP
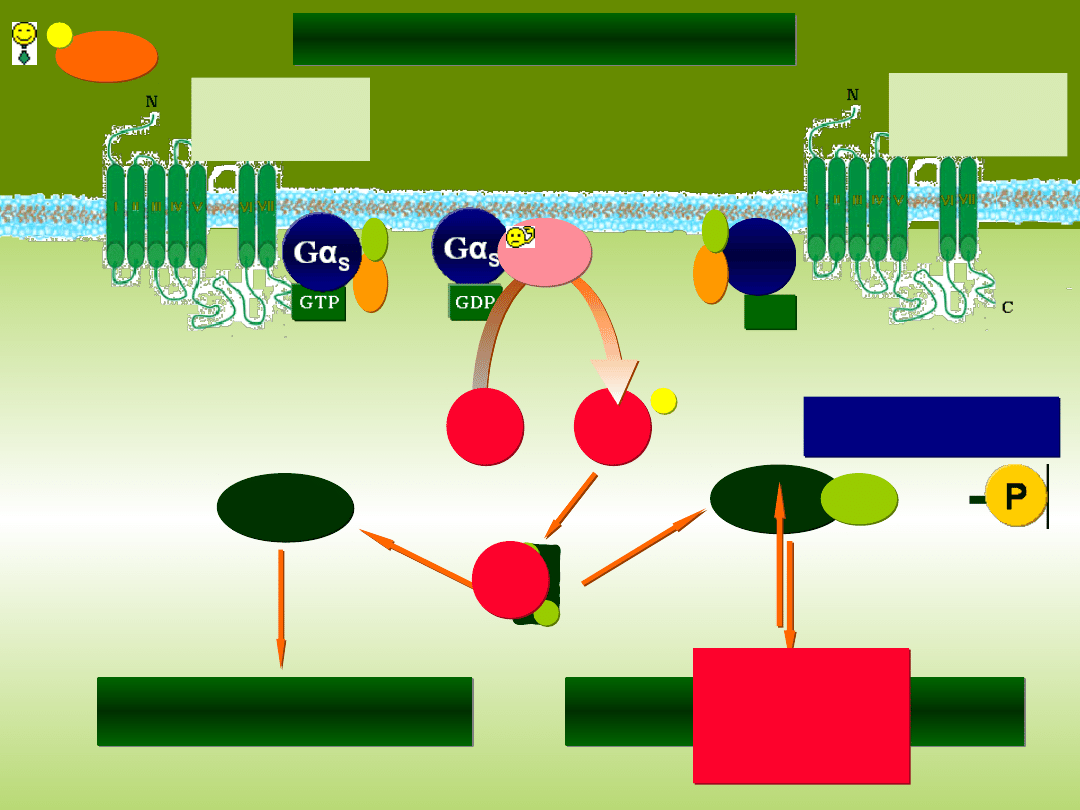
RECEPTORY METABOTROPOWE
RECEPTORY METABOTROPOWE
Gα
I
Gα
I
β
β
γ
γ
β
β
γ
γ
Gα
S
Gα
S
cAMP
cAMP
cAMP
cAMP
C
C
C
C
C
C
C
C
AC
AC
AC
AC
Białko
Białko
Białko
Białko
seryna
seryna
Zmiana aktywności enzymów
wewnątrzkomórkowych
Zmiana aktywności enzymów
wewnątrzkomórkowych
CREB
CREB
CREB
CREB
Regulacja transkrypcji
określonych genów
Regulacja transkrypcji
określonych genów
+
+
+
+
+
fosfodiesteraza
fosfodiesteraza
ATP
ATP
ATP
ATP
AMP
AMP
AMP
AMP
KOFEINA
KOFEINA
TEOFILINA
TEOFILINA
KOFEINA
KOFEINA
TEOFILINA
TEOFILINA
-
Receptor
związany z
heterotrimeryczny
m białkiem G
Receptor
związany z
heterotrimeryczny
m białkiem G
LIGAND
LIGAND
LIGAND
LIGAND
A
A
a
a
PKC
PKC
PKC
PKC
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
KALMODULINA
KALMODULINA
GDP
GDP
LIGAND
LIGAND
LIGAND
LIGAND
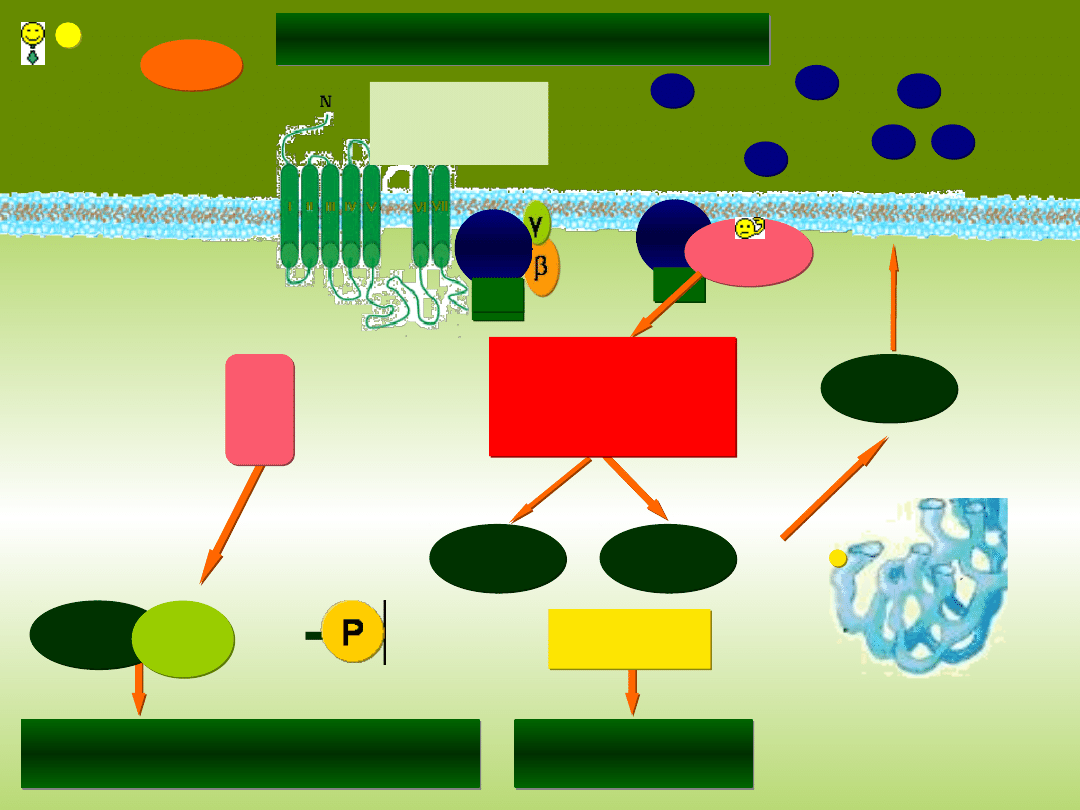
Gα
q
Gα
q
Gα
q
Gα
q
GTP
GTP
Gα
q
Gα
q
GDP
GDP
PLC
PLC
β
β
1
1
PLC
PLC
β
β
1
1
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Difosforan
Difosforan
fosfatydyloinozytolu
fosfatydyloinozytolu
PIP
PIP
2
2
Difosforan
Difosforan
fosfatydyloinozytolu
fosfatydyloinozytolu
PIP
PIP
2
2
IP
IP
3
3
IP
IP
3
3
Aktywacja kinaz
Aktywacja kinaz
IP
IP
4
4
IP
IP
4
4
SER
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Ca
2+
Białko
Białko
Białko
Białko
seryna
treonina
seryna
treonina
Zmiana aktywności enzymów
wewnątrzkomórkowych
Zmiana aktywności enzymów
wewnątrzkomórkowych
DAG
DAG
DAG
DAG
RECEPTORY METABOTROPOWE
RECEPTORY METABOTROPOWE
Receptor
związany z
heterotrimeryczny
m białkiem G
A
A
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
Wyszukiwarka
Podobne podstrony:
fizjologia-mięsnie ręki, AWF KATOWICE, FIZJOLOGIA
Wybrane zagadnienia z fizjologii mięśni i fizyczne właściwości mięśni, Biomechanika
fizjologia mięśnie
WŁAŚCIWOŚCI FIZJOLOGICZNE MIĘŚNIA SERCA, ZDROWIE, Kardiologia
Fizjologia mięśni 2
Fizjologia mięśni szkieletowych - laborki, ZDROWIE, MEDYCYNA, Anatomia i fizjologia człowieka, Anato
Fizjologia mięśni - fizjologia zwierząt. 5fantastic.pl , Ćwiczenia
39 BUDOWA I FIZJOLOGIA MIĘŚNI SZKIELETOWYCH
fizjo mail, Fizjologia mieśni gładkich, B
FIZJOLOGIA CZŁOWIEKA ĆWICZENIA, Fizjologia mini poprzecznie prkowanych, Fizjologia mięśni poprzeczni
FIZJOLOGIA - mięśnie cd, Wykłady, FIZJOLOGIA
fizjologia miesni poprzecznie prazkowanych, Studia, Neurobiologia
FIZJOLOGIA MIĘŚNI
WŁAŚCIWOŚCI FIZJOLOGICZNE MIĘŚNIA SERCOWEGO(1), BILOGIA, FIZJOLOGIA CZŁOWIEKA
Fizjologia mięśni i miomechanika, Biologia, Fizjologia zwierząt
FIZJOLOGIA MIĘŚNIA SERCOWEGO – AKTYWNOŚĆ SKURCZOWA MIĘŚNIA SERCOWEGO - instrukcja, Medycyna i Zdrowi
krótkie wykłady kraba seby z fizjologi Mięśnie, skurcz mięśnia
więcej podobnych podstron