
1
C
YTOGENETYKA
sem.
18.04.2011
Konspekt –
MECHANIZMY ABERRACJI STRUKTURALNYCH
Aberracja strukturalne:
৵
wewnątrzchromosomowe (duplikacje, delecje, inwersje)
৵
międzychromosomowe (translokacje)
৵
zrównoważone (translokacje zrównoważone, inwersje)
৵
niezrównoważone (duplikacje, delecje, chromosomy pierścieniowe,
izochromosomy, chromosomy markerowe)
Delecje:
- terminalne (pojedyncze pęknięcie w obrębie
jednego z ramion chromosomu)
/utrata części chromosmu na skutek jego
pojedynczego pęknięcia
- interstycjalne (wynik podwójnego
pęknięcia i delecji fragmentu pomiędzy
miejscami pęknięć)
mechanizm powstawania: wynik segregacji rodzinnej inwersji lub translokacji
chromosomowej; towarzysza powstawaniu izochromosomu i chromosomu
pierścieniowego
2
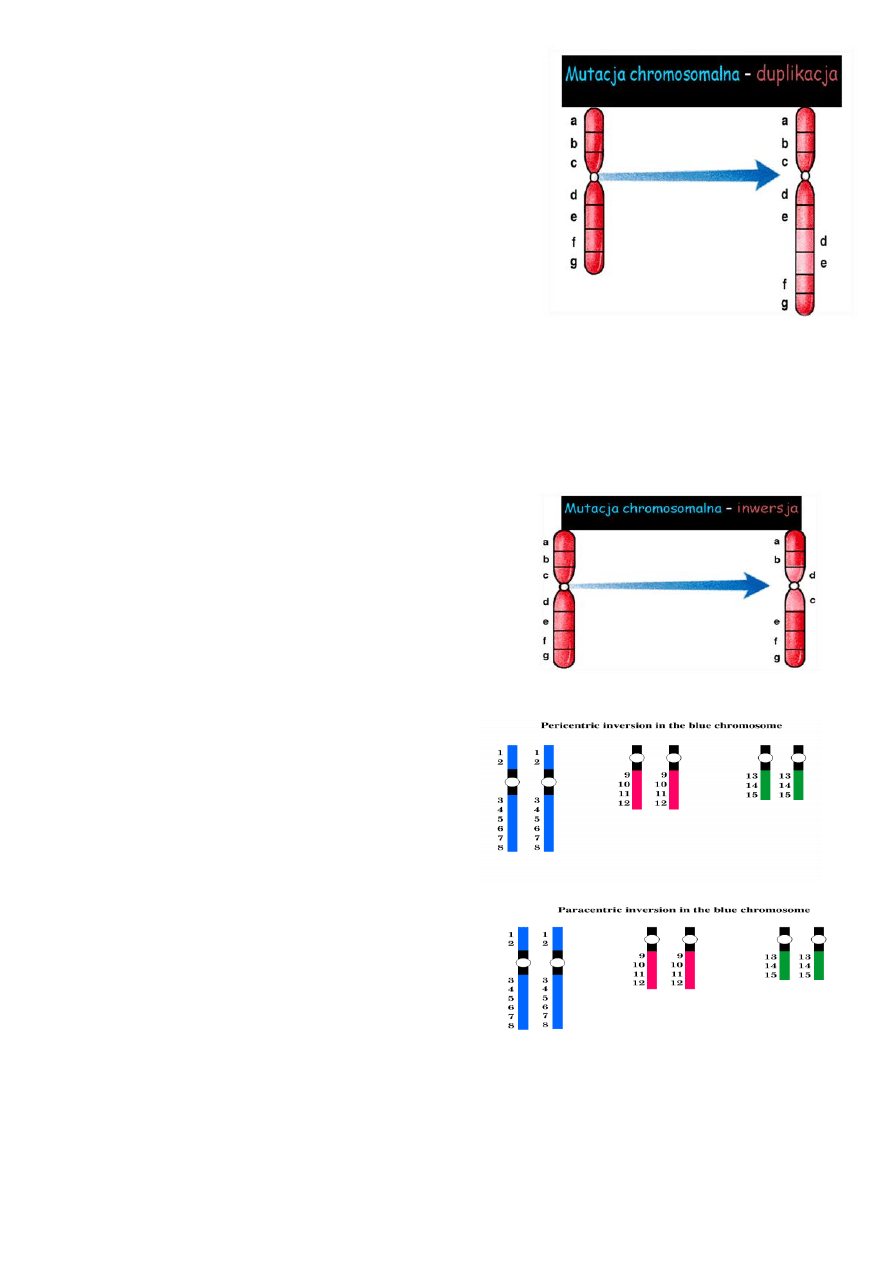
Duplikacje:
Najczęściej powstają w wyniku:
• translokacji
• inwersji
• utworzenia izochromosomu
• nierównego crossing-over podczas mejozy
Mogą występować jako:
• powtórzenia proste (duplikacje tandemowe)
• powtórzenia odwrócone względem siebie (duplikacje odwrócone)
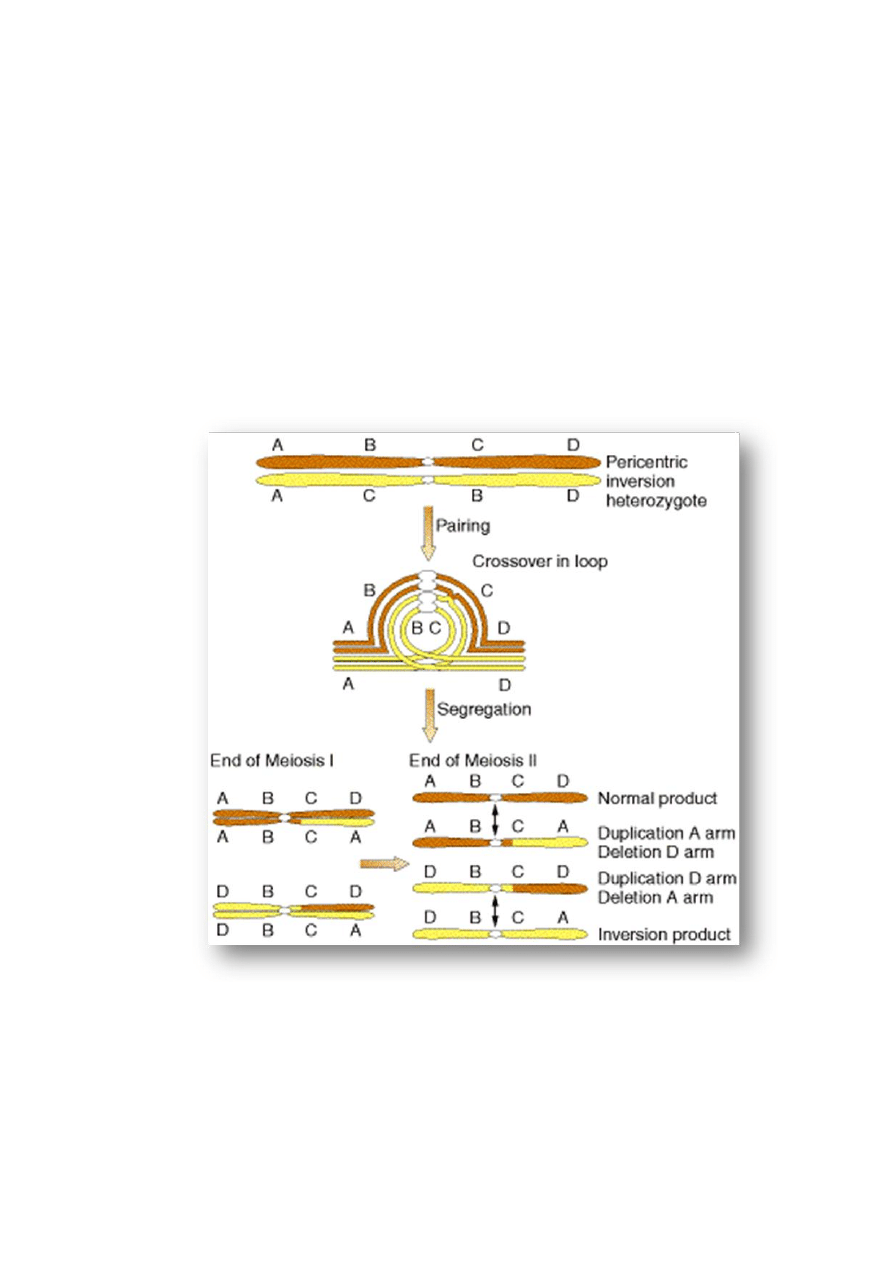
Inwersja (
odwrócenie fragmentu chromatyny między
dwoma miejscami złamań w chromosomie)
- inwersja pericentryczna (
odwrócony
fragment zawiera centromer (punkty złamań
znajdują się w obu ramionach chromosomu)
- inwersja paracentryczna (
inwersja fragmentu
jednego ramienia chromosomu)
3
Zakłócenia w podziale mejotycznym i ich następstwa:
obecność inwersji zakłóca łączenie się w pary chromosomów homologicznych podczas
mejozy; odwrócony fragment chromosomu tworzy pętlę uniemożliwiającą kontakt między
homologicznymi regionami
W przypadku
৵
inwersji pericentrycznych crossing-over w obrębie pętli prowadzi
do powstania chromosomów z
duplikacjami i delecjami dystalnych fragmentów chromosomu w stosunku do miejsc złamań w
których powstała inwersja
4
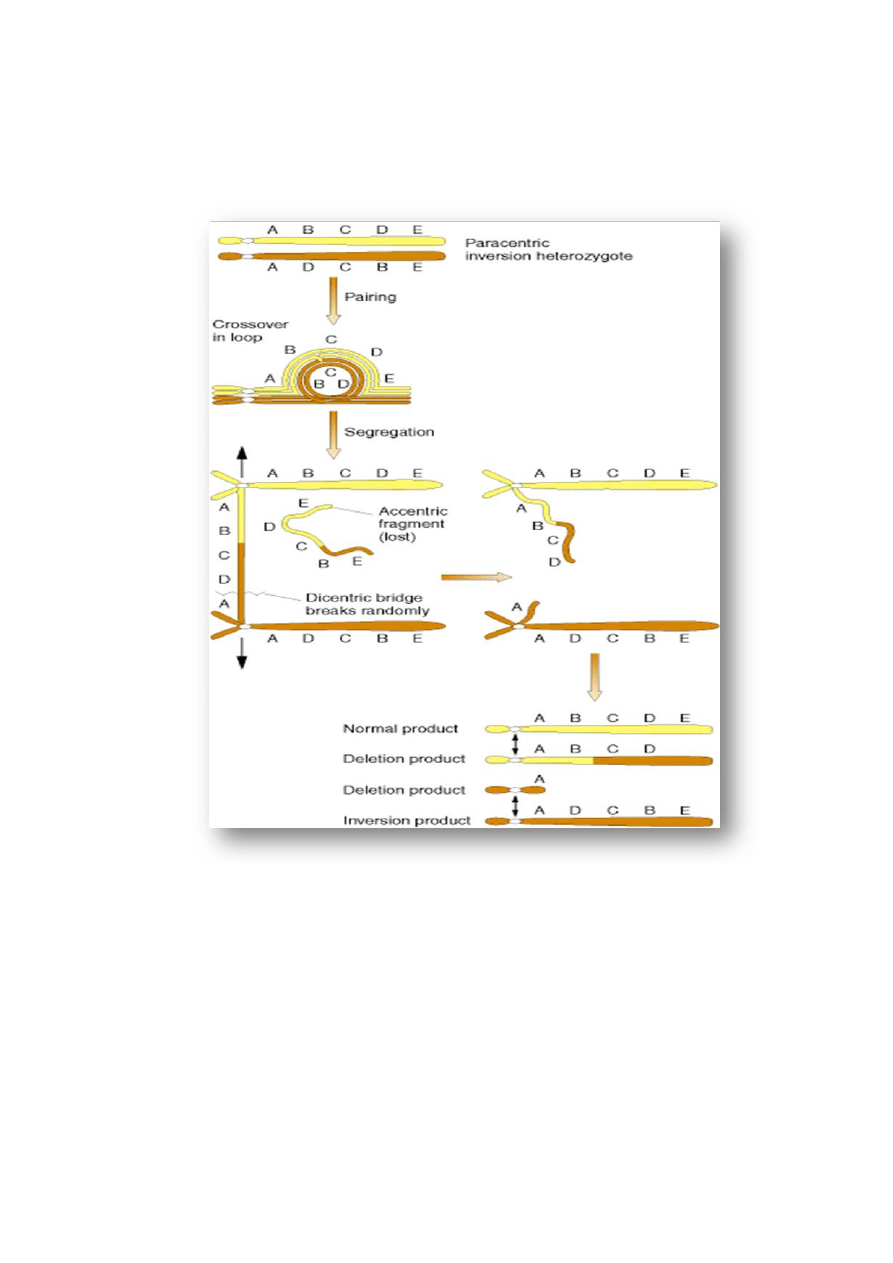
৵
inwersji paracentrycznej powstają podczas rekombinacji niestabilne chromosomy dicentryczne
i
acentryczne (z 2 centromerami lub bez centromeru) zawierające delecje i duplikacje
5
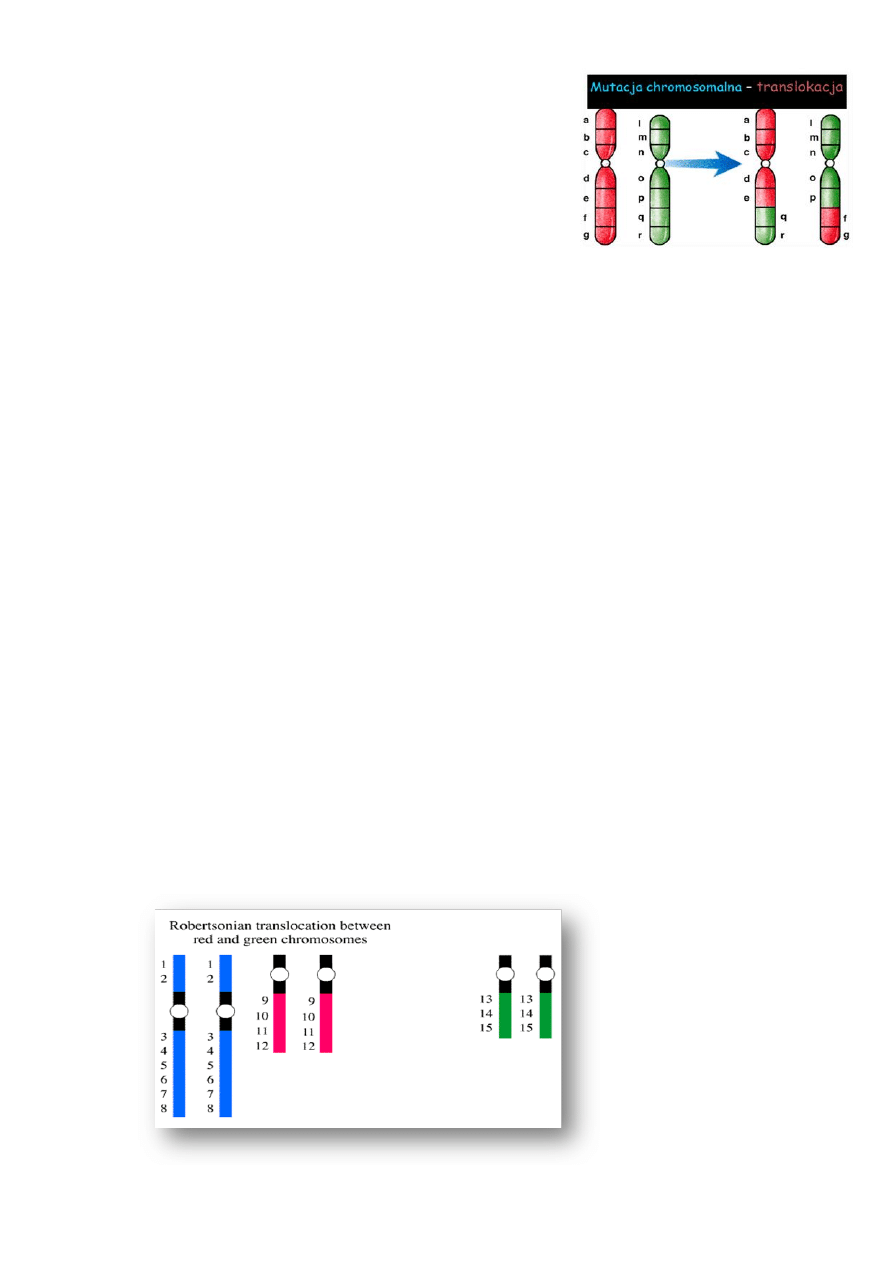
Translokacja
–
najczęściej występująca
aberracja
strukturalna
Powstają w wyniku:
- złamania chromosomów i przegrupowania fragmentów
między dwoma chromosomami
Wyróżnia się:
- translokacje wzajemne
- fuzje centryczne (translokacje - robertsonowskie)
- translokacje insercyjne
Translokacja zrównoważona -
zasadniczo nie zmienia się ilość materiału genetycznego, ale następuje
zmiana jego rozmieszczenia w genomie; liczba chromosomów może być prawidłowa lub zmieniona; aberracja
może nie przejawiać się fenotypowo
Translokacja niezrównoważona -
zmianie ulega ogólny skład genowy; ilość materiału jest większa, a liczba
chromosomów jest prawidłowa; zawsze dochodzi do ujawnienia fenotypowego choroby
Translokacja robertsonowska (fuzja centryczna) - dotyczy zawsze chromosomów akrocentrycznych
(chromosomy 13-15, 21 i 22), w których złamanie następuje w centromerze lub blisko niego; w wyniku
połączenia dwóch chromosomów akrocentrycznych powstaje jeden, często z dwoma centromerami, a krótkie
ramiona chromosomów uczestniczących w fuzji są najczęściej eliminowane; są to translokacje zrównoważone
(krótkie ramiona chromosomów akrocentrycznych są zbudowane z nieczynnej genetycznie heterochromatyny);
fuzje centryczne chromosomu 13 i 14 oraz 14 i 21 są najczęściej występującymi translokacjami u człowieka
(częstość 1/1500 osób). Segregacja chromosomów z translokacją robertsonowską podczas podziału
mejotycznego: w wyniku segregacji chromosomów mogą powstawać gamety nullisomiczne i disomiczne
względem danego chromosomu (zygoty zawierają wtedy monosomie lub trisomie całych chromosomów).
6
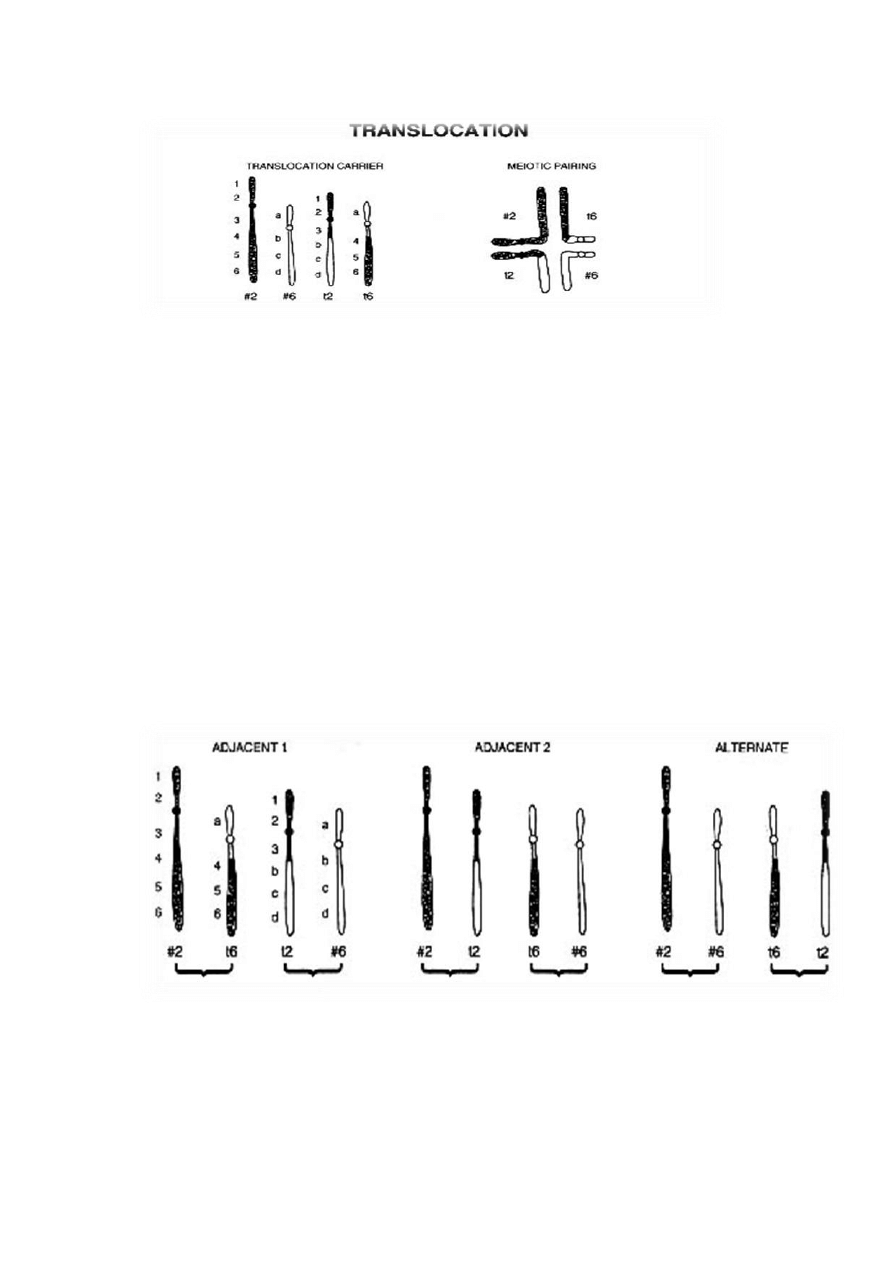
Segregacja chromosomów z translokacją wzajemną podczas podziału mejotycznego:
podczas łączenia chromosomów homologicznych w pary (Mejoza I, profaza I, zygoten) każdy z prawidłowych
chromosomów danej pary łączy się ze swoim nieprawidłowym homologiem; równocześnie homologiczne
fragmenty obu chromosomów uczestniczących w translokacji też tworzą parę – powstaje czteroramienna
konfiguracja chromosomów. Po rozdzieleniu tej struktury w anafazie do obu komórek potomnych wędrują po 2
chromosomy (segregacja 2:2) lub 3 chromosomy do jednej komórki i 1 do drugiej komórki potomnej (segregacja
1:3).
Typy rozdziału chromosomów:
[1] rozdział naprzemienny – jedna komórka potomna otrzymuje zrównoważoną translokację, druga
prawidłowe homologi chromosomów uczestniczących w translokacji
[2] i [3] rozdział przyległy typu I i II– obie gamety są nieprawidłowe i zawierają duplikacje oraz delecje
przegrupowanych fragmentów chromosomów, a zygoty mają częściowe trisomie i monosomie

7
SEGREGACJA CHROMOSOMÓW Z TRANSLOKACJĄ ROBERTSONOWSKĄ PODCZAS I
PODZIAŁU MEJOTYCZNEGO
w wyniku segregacji chromosomów mogą powstawać gamety nullisomiczne
i disomiczne względem danego chromosomu (zygoty zawierają wtedy monosomie
lub trisomie całych chromosomów)
Chromosom pierścieniowy - powstaje w wyniku połączenia dwóch złamanych końców jednego lub
kilku chromosomów, czemu towarzyszy utrata fragmentów chromosomu dystalnych w stosunku do
miejsc złamań; chromosomy pierścieniowe są niestabilne

8
Izochromosomy - chromosomy składające się z dwóch
identycznych (długich lub krótkich) ramion powstają w wyniku
duplikacji jednego i delecji drugiego ramienia chromosomu; Mogą
powstawać w wyniku nieprawidłowego, poprzecznego
(prostopadle do prawidłowej linii podziału) podziału
chromosomu w obrębie centromeru. U człowieka najczęściej
stwierdza się izochromosom długich ramion chromosomu X (w zespole Turnera). Izochromosomy
większości autosomów (poza nielicznymi wyjątkami - izochromosomy krótkich ramion
chromosomu 9, 12 i chromosomów akrocentrycznych) są letalne.
Jeśli u jednego z rodziców stwierdza się obecność izochromosomu autosomu – ryzyko wystąpienia
trisomii u potomstwa = 100%, bo w czasie mejozy izochromosom może być przekazany tylko do jednej
z komórek potomnych (powstają gamety nulli- i disomiczne)
Chromosomy markerowe - Częstość występowania: 2-7/10 000 urodzeń; Dodatkowe strukturalnie
różne chromosomy znajdowane podczas cytogenetycznych badań pre- i postnatalnych są definiowane
jako nadliczbowe chromosomy markerowe (SMCs); powstają w wyniku przegrupowań strukturalnych
materiału chromosomowego; chromosomy markerowe występujące konstytucyjnie są małe (zwykle
mniejsze od chromosomu 22); mogą to być chromosomy meta- lub akrocentryczne lub o strukturze
pierścieniowej; zawierają 1 lub 2 centromery; często wykazują obecność satelitów; mogą wywodzić się z
każdego chromosomu i mieć różny kształt; w około 60% przypadkach występowania sSMC powstają de
novo, w 40% przypadków są dziedziczone; w około 30% przypadków obserwuje się kliniczne następstwa
ich występowania

9
Wpływ obecności chromosomów markerowych na fenotyp zależy od:
- zawartości euchromatyny,
- pochodzenia chromosomowego,
- stopnia mozaikowatości
- rodzicielskiego pochodzenia
Bardzo małe SMCs zawierające materiał pochodzący z krótkich ramion chromosomów 13, 14, 15 i 21 -
niskie ryzyko nieprawidłowości klinicznych, w przeciwieństwie do wywodzących się z chromosomu 22
o
SMCs(15) stanowią ~ 50% wszystkich markerów; pacjenci nosiciele SMC(15) złożonych wyłącznie z
heterochromatyny lub/i z niewielkiej ilości euchromatyny (punkt pęknięcia: 15q11.1) są klinicznie
zdrowi; większe SMC(15) obejmujące przynajmniej region 15q12 są przyczyną licznych defektów
fenotypowych.
o
Zaledwie 15% wszystkich SMCs wywodzi się z chromosomów nieakrocentrycznych, a potencjalne
ryzyko konsekwencji klinicznych szacowane jest na 28%
o
bez wpływu na fenotyp pozostają chromosomy markerowe zawierające heterochromatynę
okołocentromerową chromosomów 1, 9 i 16, jak również te których obecność przyczynia się do
częściowych proksymalnych trisomii regionów 2q, 3p, 3q, 5q, 7p, 8p, 17p i 18p.
o
Poważne kliniczne konsekwencje towarzyszą obecności SMCs powodującej małe proksymnalne
trisomie 1p, 1q, 2p, 6p, 6q, 7q, 9p i 12q.
o
1/3 przypadków SMCs to chromosomy występujące rodzinnie. Uważa się, że takie dziedziczne markery
są nieszkodliwe, zwłaszcza jeśli są obecne u fenotypowo normalnych osób.
Miejsca łamliwe - obszary w chromosomie szczególnie podatne na pękanie chromatyd (opisanych
ponad 100)
• większość miejsc łamliwych jest związana z konkretnymi prążkami i występuje jako nieszkodliwe
warianty, z wyjątkiem fra(X)(q27.3) i fra(X)(q28), które mają związek ze specyficznymi chorobami
• wyróżniane są trzy kategorie miejsc łamliwych na podstawie ich częstości w populacji:
powszechne
o średniej częstości występowania
rzadkie (spotykane u 5% osób i w przeciwieństwie do tych częściej
spotykanych występują tylko na jednym homologu)
ZESPÓŁ ŁAMLIWEGO CHROMOSOMU X
jednostka chorobowa związana z niestabilną mutacją genu FMR-1 zlokalizowanego na
chromosomie X (mutacja dynamiczna trójki nukleotydów CGG) umiarkowany niedorozwój
umysłowy duża głowa i uszy
Wyszukiwarka
Podobne podstrony:
Konspekt mechanizmy?erracji strukturalnych (1)
Konspekt POLISACHARYDY ZAPASOWE I STRUKTURALNE PRZYKŁADY
6. konspekt - Ekonomia menedżerska, Struktury rynku a decyzje w przedsiębiorstwie
mechanizmy wytw struktur klas- warstw - 20, 20
aberracje strukturalne i genomowe chromosomów
6 konspekt Ekonomia menedżerska Struktury rynku a decyzje w przedsiębiorstwie
Referat Chemia Chemia organiczna i nieorganiczna, historia, właściwości, klasyfikacja, reakcje, mech
aberracje strukturalne i genomowe chromosomów
Wojciszke Mechanizmy wpływu struktury ja na zachowanie str 49 80
!Opis struktury - dom dziecka, Praktyka - konspekty
Konspekt LOM ZIP. s1, ZiIP Politechnika Poznańska, Obróbka Mechaniczna
Rozmieszczenie i struktura ludności Polski, Konspekty lekcji
Konspekt; rodzaje energii mechanicznej, Metodyka, Fizyka-konspekty
Temat- Parametry populacji, Konspekt lekcji Parametry populacji. Krzywa przeżywania i struktura wiek
PORÓWNANIE STRUKTURY PRZEŁOMÓW I WŁASNOŚCI MECHANICZNYCH POLIMERÓW KONSTRUKCYJNYCH
Struktura materiałów metalicznych, Akademia Morska -materiały mechaniczne, szkoła, Mega Szkoła
więcej podobnych podstron