MIKROBIOLOGIA-ZAGADNIENIA cz. II
Porównaj komórkę prokariotyczną i eukariotyczną. (ad. cz. I-13).
Opisz cykl rozwojowy u Giardia i Cryptosporidium.
Giardia Pasożyt ten jest chorobotwórczym wiciowcem należącym do rodziny Hexamitidae. Giardia intestinalis wywołuje u człowieka chorobę jelit o nazwie lamblioza (syn. giardioza) [giardiosis]. Giardia intestinalis należy do pasożytów kosmopolitycznych, to znaczy, że zarażenia tym pierwotniakiem obserwuje się na terenie całej kuli ziemskiej. Ocenia się, że w Polsce zarażonych lamblią jest od kilku do kilkunastu procent osób dorosłych. Szczególnie narażone na tego wiciowca są dzieci.
Prewalencja (częstość zarażenia) giardia intestinalis w niektórych skupiskach dziecięcych takich jak żłobki, przedszkola, domy dziecka może czasami dochodzić nawet do 100%.
Lamblia pasożytuje w organizmie człowieka w postaci 2 form, to jest cysty i trofozoitu. Trofozoit o charakterystycznym gruszkowatym kształcie jest dwubocznie symetryczny; na biegunie przednim jest zaokrąglony, natomiast w części tylnej jest zaostrzony. Na terenie cytoplazmy trofozoitu widoczne są 2 owalne jądra z chromatyną w środku oraz charakterystyczne dla lamblii kariomastigonty, aksonemy oraz ciała parabazalne. Lamblia ma również 4 pary wici: środkową, grzbietową, boczną i tylną. Na biegunie przednim trofozoitu znajduje się tak zwany krążek czepny, który umożliwia lamblii przyczepianie się do kosmków jelitowych błony śluzowej przewodu pokarmowego. W krążku czepnym znajdują się swoiste dla lamblii białka, tak zwane giardiny (α-giardina o masie cząsteczkowej 29 kDa oraz β-giardina o masie cząsteczkowej 33,8 kDa), które warunkują jego czynności fizjologiczne. Budowę trofozoitu lamblii przedstawiono na rycinie 1-6.
Cysta Giardia intestinalis jest owalna; zawiera charakterystyczną dla lamblii odstającą cytoplazmę od błony komórkowej. Na terenie cytoplazmy widoczne są 2 lub 4 jądra, aksonemy, twory sierpowate, zawiązki wici oraz liczne, bardzo drobne wodniczki jodofilne. Budowę cysty lamblii przedstawiono na rycinie 7-13.
Cysta po dostaniu się do organizmu człowieka ulega przekształceniu w procesie ekscystacjiw 2 trofozoity. Opisany proces zachodzi w dwunastnicy, gdzie trofozoity ulegają następnie dalszym podziałom, prowadząc do powstania licznej populacji trofozoitów, zdolnych do dalszej inwazji jelita cienkiego, dróg żółciowych i przewodów trzustki. W dalszych odcinkach przewodu pokarmowego trofozoity ulegają przekształceniu w procesie encystacji w cysty, które są następnie wydalane z kałem.
Do zarażenia lamblią jelitową dochodzi najczęściej drogą pokarmową przez połknięcie cyst pierwotniaka wraz z zarażoną nimi wodą lub żywnością. Możliwe jest także zarażenie lamblią poprzez stosunek seksualny, niemniej jednak do zarażenia tą drogą dochodzi niezwykle rzadko.
Giardia intestinalis wywołuje u człowieka tak zwaną lambliozę [gardiosis]. Lamblioza w pierwszym stadium chorobowym charakteryzuje się występowaniem nudności, wymiotów oraz bezkrwawej biegunki. W postaci przewlekłej tej choroby obserwuje się naprzemienne biegunki i zaparcia, bóle w nadbrzuszu, bóle głowy, stany podgorączkowe; występować mogą też objawy dyspeptyczne (wzdęcia, odbijania). Chory w tym czasie może się skarżyć na brak łaknienia oraz łatwe męczenie się. Utrzymujące się zarażenie giardia intestinalis może prowadzić do stopniowego zmniejszania się masy ciała wraz z zanikiem mięśniowym. U dzieci mogą występować choroby alergiczne, na przykład atopowe zapalenie skóry. Zauważono również, że 50% zarażeń giardia intestinalis przebiega z zaburzeniami trawienia tłuszczów, węglowodanów oraz awitaminozą A i B12; może również występować niedokrwistość niedobarwliwa, leukocytoza i eozynofilia.
Wykazano, że najbardziej podatne na zarażenie pasożytem giardia intestinalis są dzieci z niedoborami immunoglobulin klasy G i A.
Cryptosporidium parvum jest chorobotwórczym pierwotniakiem należącym do rodziny Cryptosporididae (podtyp Coccidia). Prowadzi pasożytniczy tryb życia w jelicie cienkim oraz układzie oddechowym człowieka i innych zwierząt (drób, bydło, psy, koty). Cryptosporidium parvum wywołuje u człowieka kryptosporidiozę [cryptosporidiosis] - chorobę o charakterze ostrego lub przewlekłego gastroenteritis.
Pasożyt ten z punktu widzenia medycznego ma duże znaczenie w pediatrii - atakuje szczególnie niemowlęta oraz dzieci, u których układ odpornościowy jest niedostatecznie rozwinięty i/lub wykazuje pewne dysfunkcje.
Cryptosporidium parvum jest ponadto wymieniane jako jeden z czynników tzw. biegunek podróżnych.
Podobnie jak u innych pierwotniaków należących do podtypu Coccidia (np. Isospora belli, Sarcocystis hominis, Toxoplasma gondii) cykl rozwojowy Cryptosporidium parvum jest złożony.
Wyróżnia się w nim etapy rozmnażania bezpłciowego (schizogonia) oraz etapy rozmnażania płciowego (sporogonia) - oba zachodzą na terenie organizmu żywiciela. Natomiast pewna część cyklu rozwojowego (tzn. nabywanie inwazyjności przez oocysty) zachodzi w środowisku zewnętrznym poza organizmem człowieka.
W cyklu rozwojowym Cryptosporidium parvum wyróżnia się wiele stadiów rozwojowych. Oocysta po dostaniu się do organizmu człowieka uwalnia w jelicie cienkim sporozoity, które atakują enterocyty (komórki błony śluzowej jelit), przytwierdzają się nich, a następnie przekształcają się w trofozoity. W okresie około 1 tygodnia od zarażenia trofozoity rozpoczynają etap rozmnażania bezpłciowego (schizogonię) - wytwarzane są merozoity a następnie meronty II generacji. W dalszej kolejności powstają mikro- i makrogametocyty a następnie odpowiadające im mikro- i makrogamety. Rozpoczyna się etap rozmnażania płciowego - gamety łączą się, powstaje zygota, która otacza się otoczką tworząc oocystę.
Oocysta jest następnie wydalana z kałem do środowiska zewnętrznego, gdzie staje się inwazyjna dla człowieka i zwierząt poprzez wytworzenie w jej wnętrzu czterech sporozoitów.
Opisz morfologię komórek drożdży.
Budowa komórki drożdży:
1. ściana komórkowa zewnętrzna
2. błona komórkowa (cytoplazmatyczna)
3. cytoplazma wypełniająca wnętrze komórki
4. rybosomy
5. krystaloid (jąderko)
6. jądro
7. mitochondria
8. cząsteczki białka tzw. kryształy białka
9. wodniczki
10. lipidy
Funkcje części anatomicznych komórki drożdżowej:
Ściana komórkowa - zbudowana jest z hemocelulozy, która jest substancją stosunkowo
małotrwałą w skutek czego drożdże są podatne na samounicestwienie (autoliza).
Zjawisko to polega na uszkodzeniu ściany komórkowej pod wpływem dużego stężenia enzymu zymazy przepalającego ściany komórkowe drożdży, zjawisko samounicestwienia zachodzi np. w drożdżach prasowanych piekarskich przechowywanych w niewłaściwych warunkach (wysoka temperatura i wilgotność) lub przefermentowanych rozczynach.
Błona komórkowa - zbudowana jest z zagęszczonej cytoplazmy, posiada właściwości półprzepuszczalne przenikają przez nią do wnętrza substancje odżywcze (węglowodany, tłuszcze, białka) w postaci prostej, wydzielane natomiast są na zewnątrz enzymy i produkty przemiany materii (enzymy - zymaza, oraz produkty przemiany materii - alkohol i dwutlenek węgla).
Jądro komórkowe - zawiera kwas dezoksyrybonukleinowy (DNA), którego rolą jest przekaz informacji genetycznej o budowie, składzie i funkcjach komórek potomnych.
Ponadto w skład jądra komórkowego wchodzi plazma jądrowa posiadająca charakter białkowy oraz jąderko.
Cytoplazma - jest to bezbarwna plazmoidalna ciecz o strukturze żelu, w której rozmieszczone są części anatomiczne komórki.
Substancje zapasowe i wodniczki, kropla tłuszczu, ciałko oleiste, kryształy białka - są to substancje gromadzone głównie w starszych komórkach, uzyskiwane z nadwyżek substancji odżywczych gromadzone jako materiał zapasowy posiadający charakter białkowy, tłuszczowy i węglowodanowy.
Rybosomy - odpowiadają za przekaz informacji dotyczących procesów fizjologicznych komórki takich jak oddychanie, wchłanianie przemiana materii, gospodarka enzymatyczna.
Glikogen - jest to substancja zapasowa o charakterze węglowodanowym.
Cząsteczki tłuszczu - są to substancje zapasowe składające się z tłuszczów zawierających nasycone kwasy tłuszczowe.
Klasyfikacja oraz kształty i skupiska drożdży. Kształty komórek drożdżowych: -Komórki kuliste -Komórki elipsoidalne -Komórki jajowate -Komórki mniej lub bardziej wydłużone Kształty komórek drożdży są do siebie upodobnione, w związku z czym nie mogą służyć jako cecha rozpoznawcza gatunków, kształty komórek zmieniają się wraz z wiekiem komórki i są zależne od warunków środowiska. Wielkość komórek drożdżowych: Szerokość średnio 5 μm Długość średnio 10 μm 1 μm =1 mikrometr = 0,000001цm Skupiska komórek drożdży: Drożdże tworzą skupiska w postaci łańcuszków powstałych poprzez podział komórek.
Omów sposoby rozmnażania się drożdży.
Drożdże, zarówno workowe, jak i bezworkowe, cechuje charakterystyczny sposób rozmnażania bezpłciowego w postaci tzw. pączkowania. Polega ono na tym, że w pewnym miejscu komórka tworzy rosnące stopniowo uwypuklenie, do którego przechodzi jedno z powstałych po podziale jąder potomnych. Następnie wytwarza się ściana komórkowa oddzielająca komórkę potomną od macierzystej. U drożdży piekarniczych powstałe komórki są jednakowej wielkości i oddzielają się od siebie. Po oddzieleniu, na obu komórkach (potomnej i macierzystej) pozostaje trwała blizna. W miejscu starej blizny komórka nie może utworzyć nowego pączka, a więc liczba blizn świadczy zarówno o wieku komórki, jak i o jej zdolności do rozmnażania. U grzybów drożdżoidalnych komórki często nie rozdzielają się po pączkowaniu i wydłużają tworząc tzw. pseudogrzybnię w postaci rozgałęziających się łańcuszków, gron lub krzaczków. Pseudogrzybnia (pseudomycelium) to wydłużone, nitkowate struktury, często rozgałęziające się, utworzone wyłącznie przez komórki pączkujące. Struktura ta przypomina prawdziwą grzybnię (mycelium). Ta jednak utworzona jest przez komórki dzielące się, a nie pączkujące i jest charakterystyczna dla grzybów pleśniowych.
Chociaż zdolność do pączkowania jest cechą charakterystyczną większości drożdży, to nie jest to cecha wyłącznie tej grupy grzybów (zdolność do pączkowania mają też np. niektóre podstawczaki). Niektóre drożdże nie pączkują, lecz rozmnażają się bezpłciowo przez zwykły podział komórki. Są to tzw. drożdże rozszczepkowe.
Drożdże mogą również rozmnażać się przez zarodniki, czyli spory. Oprócz wspomnianych zarodników workowych, czyli askospor (wytwarzanych tylko przez drożdże właściwe w procesie płciowym), drożdże wytwarzają też zarodniki na drodze bezpłciowej. Mogą to być blastospory, które powstają w wyniku pączkowania (charakterystyczne np. dla Candida sp.) lub chlamydospory, czyli zarodniki przetrwalnikowe, charakteryzujące się grubą ścianą.
Drożdże, czyli grzyby pączkujące zalicza się do Protoascomycetes. Typowym dla nich sposobem rozmnażania bezpłciowego jest pączkowanie. Podział komórki przez rozszczepienie występuje bardzo rzadko. Wypączkowane komórki mogą pozostawać na komórkach macierzystych i mogą tworzyć pseudo- lub pączkującą grzybnię bądź wypączkowane komórki mogą się całkowicie oddzielać. Askospory są wytwarzane w nagich workach powstałych z zygoty lub z komórek
wegetatywnych.
Grzyby z rodziny Endomycetaceae, oprócz komórek pączkujących,wytwarzają grzybnię. Endomycopsis formuje w tym samym czasie strzępki, komórki pączkujące oraz worki z zarodnikami. Endomyces lactis (znany pod nazwą Geotrichum candidum lub Oospora lactis) wytwarza artrokonidia
w wyniku fragmentacji strzępek. Saccharomycetaceae (drożdże właściwe) nie mają zdolności wytwarzania grzybni. Drożdże piwowarskie i piekarniane są odmianami fizjologicznymi Saccharomyces cerevisiae. Haploidalne komórki wegetatywne mogą się ze sobą łączyć (kopulować). Po kariogamii może następować natychmiast mejoza i formują się cztery askospory. Jednakże diploidalne komórki są zdolne do wielokrotnego rozmnażania przez pączkowanie, są one większe i bardziej aktywne fizjologicznie od komórek haploidalnych.
Drożdże nie wytwarzające worków można uznać za bardziej zredukowane (tzn. mające skrócony cykl życiowy). Tylko niektóre mają rozgałęzioną grzybnię. Większość z nich rozmnaża się wyłącznie przez pączkowanie. Drożdże nie tworzące askospor zalicza się do rodzajów:
Candida, Torulopsis, Cryptococcus, Rhodotorula, Pullularia i in. W warunkach naturalnych drożdże żyją we wszystkich siedliskach, gdzie są dostępne w postaci płynnej wydzieliny lub wydaliny zawierającej cukry łatwo podlegające fermentacji, np. w nektarze kwiatów, na owocach, na liściach. Pullularia pullulans (tzw. czarne drożdże) tworzy czarne naloty na liściach pokrytych spadzią.
Sposoby rozmnażania drożdży
przez pączkowanie np. S. cerevisiae,
przez podział np. Schizosaccharomyces pombe,
worek z zarodnikami np. S. cerevisiae,
przez pączkowanie, formująca się pseudogrzybnia np. Candida pseudotropicalis
Omów morfologię grzybów pleśniowych.
Jest to popularna nazwa określająca grzyby tworzące grzybnię zbudowaną z luźno skupionych strzępek, tzw. grzybnię powietrzną. Grzybnia ta tworzy charakterystyczne watowate skupienia o różnorodnym zabarwieniu (zielonym, zielononiebieskim, żółtym, różowym, białym lub czarnym). Wytwarza ona zarodniki i wyrasta z tzw. grzybni wegetatywnej, wrośniętej w podłoże, z którego pobiera substancje odżywcze. Podobnie jak drożdże, pleśnie również nie stanowią grupy jednorodnej systematycznie. Należą tu bowiem zarówno przedstawiciele grzybów niższych - sprzężniaki z rzędu pleśniakowców (pleśniak i rozłożek), jak i grzyby wyższe - workowce właściwe (kropidlak, pędzlak). Pleśnie rozmnażają się za pomocą zarodników.
Grzybnia wegetatywna jest plechą (łac. thallus).
Składa się ona ze strzępek o średnicy około 5 μm, wielokrotnie
rozgałęzionych i wrastających w podłoże lub rozrastających się na jegopowierzchni. Strzępki mają ścianę komórkową i cytoplazmę z organellami komórkowymi. S Sprzężniaki z rzędu pleśniakowców, jak pleśniak biały (Mucor mucedo), czy rozłożek czerniejący (Rhizopus nigricans) wytwarzają grzybnię nie podzieloną ścianami poprzecznymi (septami) na poszczególne komórki. Są więc komórczakami - tworzą jedną dużą wielojądrową komórkę. Wytwarzają one (bezpłciowo i na drodze płciowej) zarodniki wewnątrz kulistych zarodni (sporangiów) osadzonych na trzonkach zarodnionośnych (sporangioforach). Pleśnie te wytwarzają więc zarodniki wewnętrzne.
Pleśnie należące do workowców, kropidlaki (Aspergillus spp.) i pędzlaki (Penicillium spp.), tworzą grzybnie wielokomórkowe, a więc podzielone ścianami poprzecznymi i wytwarzają tzw. zarodniki konidialne (konidiospory), czyli konidia. Konidia to zarodniki tworzone zewnętrznie, tj. przez odcięcie końcowej (górnej) części specjalnej, pionowo wzniesionej strzępki, zwanej strzępką konidionośną albo konidioforem. Morfologia strzępek zarodnionośnych i zarodników pleśni jest podstawą ich identyfikacji. Cała strzępkowa plecha grzyba jest nazywana grzybnią (mycelium). W pewnych stadiach, zwykle w okresie przejścia w fazę rozmnażania płciowego lub bezpłciowego, grzybnia tworzy skupienia przypominające
tkankę nazywaną plektenchymą. Typową plektenchymą jest miąższ owocników pieczarki. U grzybów wyższych grzybnia może także formować grubsze sznury, tzw. ryzomorfy, które służą do transportu substancji odżywczych.
Omów sposoby rozmnażania się grzybów pleśniowych.
Wzrost i rozmnażanie. Strzępki grzyba wydłużają się tylko w strefie wierzchołkowej (wzrost szczytowy). U większości grzybów każda część grzybni ma potencjalną możliwość rozwoju; mały fragment grzybni jest wystarczający jako inokulum do wytworzenia nowej plechy.
Formy i mechanizmy związane z rozmnażaniem grzybów są jednak niezmiernie zróżnicowane i są one uznawane za podstawowe w klasyfikacji.
Wyróżnia się dwa typy rozmnażania, określane jako płciowe i bezpłciowe. Większość grzybów rozmnaża się oboma sposobami.
Bezpłciowo grzyby rozmnażają się głównie przez pączkowanie, fragmentację grzybni lub dzięki tworzeniu spor. Wytwarzanie spor jest najczęstszym i najbardziej zróżnicowanym sposobem reprodukcji. Konidia (zarodniki konidialne) są zwykle formowane z wierzchołkowych części strzępek powietrznych (np. u Penicillium, Aspergillus). Gdy zarodniki
tworzą się wewnątrz sporangiów (tzn. zarodni), wówczas grzyby zalicza się do sporangiosporowych (np. Mucor, Rhizopus). U grzybów niższych zarodniki często są uwicione i nazywane wtedy zoosporami lub pływkami. Ich wić ma budowę typową dla wszystkich organizmów eukariotycznych; wyrasta ona z blefaroplastu i składa się z jedenastu równoległych włókien, z których dziewięć jest położonych peryferyjnie, a dwa - centralnie (9 + 2).
Rozmnażanie płciowe. U grzybów łączenie się jąder może następować w różnym czasie od
pierwszego zetknięcia się strzępek macierzystych. W procesie płciowym można zwykle wyróżnić trzy fazy. Najpierw następuje plazmogamia, a więc łączenie się dwu protoplastów. Powstająca komórka zawiera dwa jądra. Ta para jąder, czyli dikarion, nie musi się natychmiast połączyć, a może pozostawać w stadium dikariotycznym w czasie kolejnych
podziałów komórki, oba jądra dzielą się równocześnie (podział koniugacyjny). Łączenie się dwu haploidalnych jąder (kariogamia) może następować później, często dopiero w formującym się owocniku grzyba, a powstające jądro diploidalne jest zygotą. Po kariogamii zachodzi mejoza, podział redukcyjny przywracający haploidalną liczbę chromosomów.
Te trzy procesy, czy fazy: plazmogamia, kariogamia i mejoza mogą u niektórych grzybów następować niezwłocznie jeden po drugim, u innych zaś mogą one przebiegać w zupełnie różnych stadiach rozwojowych.
U grzybów niższych proces płciowy jest zapoczątkowany przez tworzenie gamet — komórek płciowych. Gdy gamety formujące się w męskich i żeńskich gametangiach są morfologicznie nierozróżnialne, nazywamy je izogametami. Gamety często tworzą się wewnątrz gametangiów, które różnią się od siebie, wtedy męskie gametangium jest nazywane
anteridium (plemnia), a żeńskie oogonium (lęgnia). Gdy męskie i żeńskie gametangia tworzą się na tej samej grzybni wegetatywnej (powstałej z jednego zarodnika), grzyb jest uznawany za homotalliczny, czyli hermafrodytyczny (monozoiczny). Grzyby heterotalliczne, czyli dizoiczne, mają plechy męskie lub żeńskie. Grzyby homotalliczne mogą być samozgodne płciowo (autogamiczne). Jednak u niektórych grzybów homotallicznych następuje samozapłodnienie z powodu działania fizjologicznego inhibitora, który jest odpowiedzialny za samo niezgodność (inkompatybilność). To sprawia, że np. u Neurospora, chociaż jest
homotalliczna, przy zapładnianiu niezbędne jest połączenie (koniugacja) dwu różnych typów plech (+ i —); plechy należące do tego samego typu są niezgodne płciowo.
Na czym polega fotosynteza oksygenowa i jakie organizmy ją przeprowadzają?
Przeprowadzają ją glony i sinice.
Podstawowe procesy fotosyntezy odbywają się w tylakoidach. Są to spłaszczone, całkowicie zamknięte pęcherzyki błonowe występujące w komórkach sinic oraz w chloroplastach zielenic i roślin wyższych.
Błony tylakoidowe w chloroplastach zawierają chlorofil (Chl) a i Chl b, karotenoidy, przenośniki elektronóworaz enzymy. Większość cząsteczek chlorofilu (99,5%) oraz pomocniczych barwników (karotenoidy, fikobiliny) uczestniczy w absorpcji światła.
W fotosyntezie oksygenowej występują szeregowo dwa systemy pigmentów (PS I i PS II) i przekazie energii. Tworzą one system barwników anten. Obydwa fotosystemy pigmentów są połączone łańcuchem transportu elektronów i współpracują ze sobą.
Na czym polega fotosynteza anoksygenowa i jakie organizmy ją przeprowadzają?
Przeprowadzają ją bakterie zielone i purpurowe.
W fotosyntezie anoksygenowej bierze udział pojedyncza reakcja świetlna, która napędza cykliczny transport elektronów. Elektrony wycofane z cyklu do redukcji NAD nie zostają odtworzone przez rozszczepienie wody. Fotosynteza jest zatem zależna od obecności w podłożu zredukowanych substancji i przebiega bez wydzielania tlenu. Chociaż przebieg właściwej reakcji świetlnej jest analogiczny do takiej reakcji u roślin, to prawdopodobnie prowadzi ona jedynie do powstania siły protonomotorycznej, a zatem do wytworzenia energii (ATP), lecz nie do redukcji NAD
Obydwa fotosystemy pigmentów są połączone łańcuchem transportu elektronów i współpracują ze sobą.
Porównaj fotosyntezę oksy- i anoksygenową.
Jeśli chodzi o zamianę energii świetlnej w energię wykorzystywaną w procesach biochemicznych (ATP),to w zasadzie nie ma większych różnic między fotosyntezą bakterii fototroficznych i roślin zielonych. U bakterii purpurowych fotosynteza wydaje się ledwo wystarczać do pokrycia ilości wykorzystanej energii. Fotosynteza u sinic i roślin zielonych pod względem ewolucyjnym jest procesem bardziej rozwiniętym; ciąg dwóch kolejnych reakcji świetlnych umożliwia aktywację elektronów w pierwszej reakcji do poziomu wystarczającego do redukcji ferredoksyny i NADP. Druga reakcja świetlna pozwala następnie na wykorzystanie wody jako źródła elektronów. Poza uzyskiwaniem energii, układ ten umożliwia również redukcję NADPi wydzielanie tlenu.
Scharakteryzuj purpurowe bakterie siarkowe.
Purpurowe bakterie siarkowe - są to bakterie, które w swoim metabolizmie wykorzystują mechanizm, jakim jest utlenianie siarki (tyczy się to wszystkich bakterii siarkowych). Nazywane Protobakterie. Jest to największa i najbardziej zróżnicowana grupa prokariotów. Te bakterie gromadzą siarkę w postaci silnie załamujących światło kuleczek. Są to beztlenowce, fototrofy - bo ich źródłem energii jest światło, do tego węgiel pozyskują z CO2 bo są autotrofami. Prowadzą fotosyntezę lecz beztlenową. Fotosynteza anoksygenowa. Przykłady : Chromatium. Nie są one zdolne wykorzystywać wodę jako donora wodoru więc korzystać mogą np. z siarkowodoru. Większe mają kształt fasolowaty a mniejsze to krótkie pałeczki. Kilka gatunków zawiera wakuole gazowe. Typowa cecha jednej z rodzin czyli Chromatiaceae to odkładanie siarki wewnątrzkomórkowe w trakcie utleniania siarkowodoru. Ectothiorhodospira przedstawiciele tego rodzaju akumulują siarkę na zewnątrz. Większość z bakterii purpurowych siarkowych zawiera pęcherzykowate tylakoidy (są drobne wyjątki).
Scharakteryzuj purpurowe bakterie bezsiarkowe.
Purpurowe bakterie bezsiarkowe - zalicza się je do 5 rodzajów w zależności od kształtu: spiralne, pałeczkowate, zakrzywione pałeczki, zbliżone do kulistych, ogólnie posiadają wszystkie znane struktury tylakoidalne. Dwa gatunki wyróżniają sięszczególnie. Otóż Rhodomicrobium vannieli i Rhodocyclus purpureus. Pierwszy rozmnaża się przez pączkowanie, pączki pozostają połączone z komórką macierzystą przez łodyżki podobne do strzępek lub uwalniają sięw postaci perytrychalnie urzęsionych ruchliwych komórek. A Ten drugi rodzaj to jedyna forma z tej rodziny która jest nieruchliwa. Jego komórki są półokrągłe błona fotosyntetyzująca najprawdopodobniej podobna do plazmatycznej tylko silnie pofałdowana.
Wzrost większości bakterii bezsiarkowych hamowany jest przez siarkowodór choć niektóre tolerują ten związek.
Scharakteryzuj zielone bakterie siarkowe.
Zielone bakterie siarkowe - w bezpośrednim sąsiedztwie błony cytoplazmatycznej mają organelle zawierające pigmenty (barwniki) - organelle te to tzw. Chlorosomy znane jako pęcherzyki Chlorobium. Zawierają one bakteriochlorofil c,d lub e, czyli pigment anten. Oraz zawierają niewielkie ilości bakteriochlorofilu a. Od bakterii purpurowych różni te bakterie brak enzymu karboksylazy rybulozobisfosforanowej - nie wiążą zatem CO2. Zaliczamy do tego typu bakterii dwie rodziny, które różnią się od siebie. Chlorobiaceae gramujemne i Chloroflexaceae gramdodatnie.
Te pierwsze mają zielone lub brązowe zabarwienie i tworzą skupiska przypominające gwiazdę lub sieć.
Bakterie drugiej grupy to fototrofy są to bakterie nitkowate o ruchu ślizgowym. Zawiera bakteriochlorofil c i a oraz chlorosomy podobne do chlorosomów gatunku Chlorobium. Z drugiej strony różni się tych wymienionych przed chwilą tym, że ma zdolność do heterotroficznego wzrostu na złożonych podłożach zarówno w ciemności i w świetle. Prawie niewykorzystuje H2S i CO2. Jest szeroko rozpowszechniony na całym świecie żyje na dnach kanałów (zielone i pomarańczowe kolory) i w wyciekach z gorących źródeł.
Scharakteryzuj zielone bakterie bezsiarkowe.
Bakterie z grupy Chlorofexaceae. Zawierają struktury podobne do chlorosomów i bakterio chlorofil a i c. Są tlenowcami. CO2 nie wiążą w cyklu Calvina, lecz w szlaku hydroksypropionowym. Jako źródło elektronów do redukcji NAD wykorzystują związki organiczne, rzadko H2S lub H2. Potrafią żyć bez światła i odżywiać się heterotroficznie. Mają postać nitkowatą i często tworzą maty.
Charakterystyka Halobacterium halobium.
Nie jest autotrofem - odżywia się heterotroficznie, a energia uzyskiwana z fotofosforylacji uzupełnia energię otrzymywana z oddychania tlenowego.
Na czym polega fotosynteza bakteryjna i jakie ma znaczenie w przyrodzie?
Bakterie purpurowe tworzą odmienny typ metaboliczny: purpurowe bakterie siarkowe i bezsiarkowe na świetle asymilują, odpowiednio: dwutlenek węgla oraz substancje organiczne. Ten typ fotosyntezy odbiega jednak zasadniczo od fotosyntezy roślin wyższych. Po pierwsze, nie wykorzystuje wody jako donora wodoru dla fotosyntezy, nie powstaje zatem wolny tlen. Po drugie, zużywa siarkowodór lub substancje organiczne jako donory wodoru (czego nie potrafią rośliny zielone). W wyniku dalszych badań nad bakteriami purpurowymi. W fotosyntezie bakteryjnej siarkowodór odgrywa taką rolę, jak
woda w fotosyntezie roślin wyższych.
Na czym polega chemosynteza i jakie ma znaczenie dla człowieka? Chemosynteza- biochemiczny proces przyswajania analogiczny do fotosyntezy, lecz zachodzący bez udziału światła, kosztem energii chemicznej uzyskiwanej z procesu utleniania tlenem atmosferycznym . prostych substancji nieorganicznych, jak wodór, siarka, siarkowodór, amoniak, lub organicznych, jak metan; uzyskana w tych procesach energia pozwala organizmom na związanie dwutlenku węgla i wody, i syntetyzowanie z nich własnych związków organicznych; do chemosyntezy są zdolne nieliczne gat. bakterii, zw. chemoautotrofami, z których najważniejsze to: 1) bakterie wodorowe, utleniające wodór cząsteczkowy do wody, 2) bakterie siarkowe, utleniające siarkowodór, siarkę i tiosiarczany do siarczanów, 3) bakterie nitryfikacyjne, utleniające amoniak, sole amonowe i azotyny do azotanów (nitryfikacja), 4) bakterie żelazowe, utleniające jony żelaza Fe2+ do jonów Fe3+. Proces chemosyntezy odgrywa ważną rolę w krążeniu pierwiastków w przyrodzie, a także w tworzeniu się niektórych złóż, np. azotanów (saletry), rud żelaza, manganu. Można ją podzielić na dwa etapy: 1.Utlenienie związku nieorganicznego i uzyskanie energii związek nieorg. zredukowany+tlen → związ. Nieorg. Utleniony+ energia ATP, NADPH2 2.Synteza związków organicznych:
CO2+H2O+Energia → glukoza + tlen Udział w krążeniu pierwiastków w biosferze (S, N, C, Fe), Usuwanie toksycznego H2S i NH3,Produkcja materii organicznej bez udziału światła, Udział w procesie glebotwórczym i tworzeniu złóż
Znaczenie dla człowieka:
1. wykorzystanie w procesie oczyszczania ścieków
(nitryfikacja i anamoks, metanogeneza),
2. wykorzystanie w procesie bioługowania
(chemosynteza siarkowa i żelazowa),
3. znaczenie negatywne: korozja, wytrącanie Fe III
Na czym polega chemosynteza siarkowa i żelazowa, jakie mikroorganizmy ją przeprowadzają?
2H2S + O2 → 2S +2H2O + energia ogólne równanie reakcji chemosyntezy siarkowej
Chemosynteza siarkowa polega na uzyskiwaniu związków organicznych z utleniania różnych związków siarki, np. siarkowodoru. Powstająca wolna siarka jest wydzielana na zewnątrz lub gromadzona w komórce. Siarkowe bakterie występują pospolicie na całej kuli ziemskiej, ich działalność przyczyniła się do powstania złóż siarki.
Mikroorganizmy: Thiobacillus , bakterie z rodzaju Beggiatoa , bakterie z rodzaju Thiotrix
Analogicznie chemosynteza żelazowa polega na tworzeniu związków ogranicznych z utleniania związków żelaza.
4 Fe2+ + 4H++ O2 → 4Fe3+ + 2H2O + energia
Bakterie - Ferrobacillus, Galionella, Thiobacillus
Na czym polega nitryfikacja i jakie mikroorganizmy ją przeprowadzają?
Nitryfikacja to proces biologicznego utleniania amoniaku do azotanów zachodzący przy
udziale bakterii nitryfikacyjnych. Nitryfikacja to przekształcenie NH4 + i NO2 - do NO3
-prowadzone głównie przez chemolitotrofy. Uwolniona w tym procesie energia jest wykorzystywana do syntezy związków organicznych.
Nitryfikacja przebiega dwuetapowo:
● Najpierw amoniak jest utleniany do azotynu, bakterie utleniające NH4 do NO2
określane są przedrostkiem „nitroso-”. Należą tu m.in.: Nitrosomonas, Nitrosospira,
Nitrosocyjastis, Nitrosoglea. Są to bakterie właściwe, mają kształt drobnych pałeczek.
● Następnie powstały azotyn jest utleniany do azotanu, azotyny do azotanów utlenia
grupa „nitro”: np. rodzaj Nitrobacter, Nitrospira, Nitrococcus.
● Obie grupy bakterii nitryfikacyjnych są wrażliwe na zakwaszenie środowiska;
zahamowanie ich wzrostu następuje przy pH 5.0.
Proces nitryfikacji może być prowadzony także przez mikroorganizmy heterotroficzne.
Największą grupą mikroorganizmów przeprowadzających heterotroficzną nitryfikację są
grzyby:Aspergillus flavus, Penicillium, Cephalosporium. Nitryfikacja prowadzona przez
grzyby jest mniej wrażliwa na zakwaszenie i bardziej odporna na suszę.
Wytworzone w glebie azotany mogą być przyswajane przez rośliny, wypłukiwane przez
wodę lub rozkładane w procesie denitryfikacji.
Na czym polega reakcja anamoks i jakie mikroorganizmy ją przeprowadzają?
Anamoks - proces beztlenowego utleniania amoniaku przeprowadzany przez bakterie.
preferowanymi akceptorami elektronów są azotyny (azotany (III)), a nie azotany (V) i że reakcja przebiega w następujący sposób:
NH4+ + NO2- → N2 + 2 H2O ΔG = -358 kJ/mol NH4+
Produktami pośrednimi w tym procesie są hydroksylamina i hydrazyna. Jednym z głównych enzymów biorących udział w reakcji anammox jest oksydoreduktaza hydroksylaminy, która prowadzi utlenianie zarówno hydroksylaminy, jak i hydrazyny. Enzym ten znajduje się w organellum zwanym anamoksosomem.
Bakterie przeprowadzające reakcje anammox należą do 3 rodzajów:
- brocadia
- kuenenia
- scalindua
Aktywność bakterii prowadzących tę reakcję jest 25 razy większa niż tlenowych nitrozobakterii utleniających amon w warunkach denitryfikacyjnych (beztlenowych) i 7 razy mniejsza niż aktywność nitrozobakterii utleniających amon w warunkach tlenowych.
Omów rodzaje rozmnażania u bakterii.
Rozmnażanie bezpłciowe
Bakterie rozmnażają się przez prosty podział komórki (rozszczepianie), przy czym z jednej komórki macierzystej powstają, po wytworzeniu poprzecznej błony, dwie komórki potomne. Pierwszy etap to podział substancji jądrowej. Występuje on w fazie intensywnego wzrostu komórki. Kolisty chromosom bakteryjny ulega w wyniku replikacji podwojeniu. Po skończonym podziale nukleoidu następuje właściwy podział komórki bakteryjnej, tworzy się poprzeczna przegroda (septum) rosnąca od zewnątrz do środka komórki. Stanowi ona następnie ścianę komórkową odgradzającą nowo powstałe komórki. Istotny jest brak wrzeciona kariokinetycznego, jakie podczas mitozy tworzy się u Eukaryota.
U bakterii nie stwierdzono rozmnażania płciowego, wykryto natomiast procesy płciowe umożliwiające pewną wymianę materiału genetycznego między różnymi komórkami bakteryjnymi.
1. Koniugacja:
Przenoszenie dziedzicznych cech szczepu dawcy na szczep biorcy przez bezpośredni kontakt w parach określa się mianem koniugacji. Połączenie się 2 bakterii zachodzi poprzez mostek cytoplazmatyczny utworzony przez pili płciowe. Koniugują tylko bakterie F+ (dawca, zawiera czynnik płciowy F i może wytwarzać pili płciowe) z bakterią F- (biorca, nie ma czynnika F). Podczas koniugacji biorca może uzyskać czynnik F i zacznie wytwarzać pili płciowe, stanie się dawcą.
2. Transformacja:
Przekazanie cech genetycznych szczepom biorcy z pominięciem łączenia w pary, za pomocą wolnego rozpuszczalnego DNA uzyskanego od dawcy.
3. Transdukcja:
Jest to proces przenoszenia fragmentu DNA z jednej komórki do drugiej przez bakteriofaga łagodnego (w czasie cyklu lizogenicznego). Fag taki wbudowuje część bakteryjnego DNA w swój własny kwas nukleinowy i przenosi go jako trwałą cechę na szczep biorcy.
Omów cykl rozwojowy Chlamydia pneumoniae, Bdelliovibrio sp. i bakterii śluzowych.
Bakterie śluzowe
Bakterie śluzowe to ścisłe tlenowce chemoheterotroficzne, zdolne do ruchu pełzającego
Cykl rozwojowy z wytworzeniem ciał owocowych:
Po wytworzeniu skupiska komórek powstaje ciało owocowe, składające się ze śluzowej nóżki i z cyst. Cysty są organami do rozprzestrzeniania i w czasie kiełkowania uwalniaja mikrospory, które rozwijają się w komórki wegetatywne.
Bdellovibrio
Jest tlenowym mikroorganizmem pasożytującym na innych bakteriach Napotkawszy na odpowiednią bakterie żywiciela pasożyt ten przywiera nieurzęsionym biegunem do jej ściany komórkowej i obraca się wokół swej osi podłużnej. Wkrótce potem zaatakowana komórka przekształca się w formę kulistą ,podobną do sferoplastu. Bdellovibrio penetruje jej ścianę komórkową i umiejscawia się w przestrzeni peryplazmatycznej . Komórka rośnie i wydłuża się, aż do chwili kiedy zostaną wyczerpane związki pokarmowe ze stopniowo malejącego protoplastu żywiciela. Forma cylindryczna dzieli się wielokrotnie, dając komórki o jednakowych rozmiarach. W końcu ściana komórkowa gospodarza ulega lizie i do podłoża zostaje uwolnione potomstwo pasożyta gotowe do zaatakowania nastepnych bakterii.
Chlamydia pneumoniae:
Chlamydie są patogenami ludzi,na podstawie cech biologicznych zalicza się je do prokariotów. Syntetyzują związki, których nie potrafią syntetyzować komórki eukariotyczne, takie jak kwas muraminowy, kwas foliowy i kw. diaminopimelinowy.
Wszystkie bakterie z tej grupy mają podobny cykl reprodukcji. Ciałka elementarne (EB elementary bodies), malutkie komórki o średnicy około 300 nm. Zawierają nukleoid. Są to formy inwazyjne dla komórek, do których wnikają w wyniku fagocytozy. Po 6-8 godzinach przekształca się w ciałko pierwotne o rozmiarach od 0.5 do 1 μm, które pozbawione jest nukleoidu (pozostaje wodniczka) i błon powierzchniowych z komórki gospodarza. Wewnątrz znajduje się ciałko RB (reticulate body). W ciągu następnych 18-24 godzin ciałko powiększa się,a zawarte RB ulegają wielokrotnym podziałom. Otaczająca je wodniczka wypełnia się ciałkami wtrętowymi. Następnie ciałka są uwalniane poza komórkę i już jako ciałka EB zakażają kolejne komórki powtarzając cykl.
Co to są formy L u bakterii i formy pleomorficzne?
Podział komórki bakteryjnej i wzrost populacji bakterii w czasie.
Hodowla okresowa-charakterystyka ogólna.
W hodowli statycznej (okresowej) mikroorganizmy posiane do pożywki rosną i rozmnażają się do czasu wyczerpania się składników pokarmowych lub (i) nagromadzenia się toksycznych produktów przemiany materii. W tego typu hodowli rozwój populacji bakterii przebiega w kilku charakterystycznych fazach, które można zobrazować na wykresie w postaci tzw. krzywej wzrostu:
faza zastoju,
faza wzrostu logarytmicznego (wykładniczego),
faza stacjonarna,
faza zamierania.
W fazie zastoju w zaszczepionych komórkach (inokulum) zachodzą procesy adaptacji polegające na syntezie potrzebnych enzymów, replikacji DNA, syntezie białek i w efekcie komórki zwiększają swoje rozmiary. Długość tej fazy zależy od podobieństwa warunków hodowli poprzedniej (z której pochodzi inokulum) do warunków panujących w nowej hodowli. Im jest ono większe, tym faza jest krótsza.
W fazie wzrostu logarytmicznego komórki zaczynają się dzielić. Sygnałem do podziałów jest osiągnięcie przez komórki odpowiedniej długości. Każda komórka dzieli się na dwie. Po określonym czasie wzrostu powstałe komórki znowu dzielą się na dwie, stąd liczbę powstałych komórek (czyli wzrost populacji) określa wzór 2n, gdzie n - to liczba podziałów, która jest równoznaczna z liczbą pokoleń. Czas między dwoma kolejnymi podziałami, to tzw. czas generacji lub wiek osobniczy. Zależy on od warunków hodowli i od cech gatunkowych drobnoustroju. W konkretnej hodowli jest on więc stały. Jeśli liczba komórek w inokulum wynosi N0, to powstała liczba komórek N po n pokoleniach wyniesie N = N0 x 2n. Liczba bakterii podwaja się co każdy okres generacji, rośnie więc wykładniczo z upływem czasu. Do czasu hodowli proporcjonalny jest więc logarytm liczby bakterii, a nie sama ich liczba. Stąd nazwa - faza logarytmiczna.
W fazie stacjonarnej obserwuje się spadek przyrostu liczby bakterii, w wyniku zamierania części komórek z powodu wyczerpywania się składników pokarmowych, tlenu i wytwarzania produktów przemiany materii. Zamieranie to jest w pewnej równowadze z dzieleniem się innych komórek
Z czasem komórek zamierających jest więcej i dochodzi do spadku ogólnej liczby komórek - hodowla się przerzedza i z czasem zamiera.
Wyjaśnij pojęcie wydajności (plonu hodowli)
Wydajnością, plonem lub uzyskiem nazywa się masę bakteryjną wytworzoną w chwili osiągnięcia fazy stacjonarnej (jedna z faz wzrostu bakterii w hodowlach okresowych, która rozpoczyna się w momencie gdy komórki mogą już się reprodukować). Zależy to od rodzaju składników odżywczych pożywki oraz warunków hodowli. Wydajność jest różnicą między początkową a maksymalną masą bakteryjną: X=Xmax - X0. Wartość ta jest wyrażana w gramach suchej masy.
Wyjaśnij pojęcie swoistej szybkości wzrostu bakterii.
Jest to miara szybkości wzrostu komórek. Oblicza się ją z gęstości bakterii x0 i xt, odpowiednio w czasie t0 i czasie t.
Hodowla ciągła-charakterystyka.
…Jednak opisana sekwencja zdarzeń w hodowli może być inna. Jeśli tylko zapewni się usuwanie zużytego podłoża i zastępowanie go świeżym, to możliwe jest utrzymywanie fazy wzrostu logarytmicznego praktycznie w nieskończoność. Na tym właśnie polega hodowla ciągła. Jest więc ona, w przeciwieństwie do hodowli statycznej, układem otwartym, z ciągłym przepływem pożywki (zużytej i świeżej). Prowadzi się ją w tzw. chemostatach, umożliwiających kontrolowanie wzrostu komórek za pomocą dozowania odpowiednich ilości pożywki i regulowania szybkości przepływu. Dzięki temu możliwe jest uzyskanie stanu równowagi, w którym zagęszczenie komórek jest stałe. Ma to duże znaczenie w badaniach procesów fizjologicznych drobnoustrojów, kiedy niezbędne są warunki stabilne. Hodowle ciągłe są wykorzystywane m.in. w przemyśle spożywczym i farmaceutycznym, do produkcji określonych substancji (np. kwasu octowego, antybiotyków) i biomasy (np. drożdży). Hodowle w procesach produkcyjnych prowadzi się w bioreaktorach (fermentorach). Rodzajem hodowli ciągłej jest również biologiczna oczyszczalnia ścieków, w której hoduje się bakterie (jako tzw. osad czynny) na bazie pożywki, jaką stanowią ciągle dopływające ścieki.
Hodowla zsynchronizowana-charakterystyka.
Hodowla zsynchronizowana to hodowla, w której komórki dzielą się równocześnie (czyli synchronicznie), w przeciwieństwie do innych hodowli, gdzie podziały komórek są nieskoordynowane. Dzięki jednoczesności podziałów, zmiany w hodowli synchronicznej są odzwierciedleniem (zwielokrotnieniem) zmian w pojedynczej komórce. Umożliwia to badanie przemian zachodzących w cyklu życiowym komórki, zwykle nieuchwytnych ze względu na zbyt małą czułość metod badawczych. Zsynchronizowanie podziałów można osiągnąć wieloma metodami, m.in. przez szok wywołany niską temperaturą. Bakterie przenosi się na ok. 15 - 60 min. do temperatury dużo niższej od optymalnej. W tych warunkach dochodzi do zrównania stanu fizjologicznego komórek, które przeniesione do temperatury optymalnej, równocześnie zaczynają podziały komórkowe. Osiągnięta synchronizacja nie jest trwała i, jeśli nie jest utrzymywana, szybko spontanicznie zanika.
Omów skład, wielkość i zdolność penetracji dróg oddechowych aerozolu biologicznego.
Mikroorganizmy w powietrzu tworzą układ koloidalny w postaci aerozolu biologicznego (bioaerozolu). W przypadku aerozoli biologicznych ośrodkiem dyspersyjnym jest powietrze (lub inny gaz), a fazą rozproszoną - mikroorganizmy.
Bioaerozol może mieć postać dwóch rodzajów faz:
fazy pyłowej (np. powstającej dzięki ruchom powietrza unoszącym kurz)
fazy kropelkowej (np. powstającej w wyniku kondensacji pary wodnej lub w czasie kichania)
Wielkość cząstek bioaerozolu
Średnica cząstek obejmuje zakres
od ok. 0,02µm do 100µm.
Stosując kryterium wielkości można podzielić aerozol biologiczny na:
drobnoziarnisty (poniżej 1µm)
gruboziarnisty (powyżej 1µm)
Wpływ wielkości cząstek bioaerozolu na penetrację dróg oddechowych:
Bioaerozol gruboziarnisty osadza się głównie w jamie nosowo-gardłowej (szczególnie cząstki o średnicy powyżej 10µm) i drzewie oskrzelowym, tzn. tchawicy, oskrzelach i oskrzelikach (cząstki o średnicy 2 - 10µm)
Bioaerozol drobnoziarnisty przenika głębiej, aż do pęcherzyków płucnych (cząstki o średnicy 1 µm i mniejszej)
Frakcjia respirabilna (łac. respirare - oddychać) to cząstki, które mogą przenikać w głąb płuc (do oskrzelików i pęcherzyków płucnych)
Opisz mechanizmy obronne organizmu człowieka przed wnikaniem bioaerozolu.
Mechanizmy usuwające bioaerozol z wdychanego powietrza:
-aparat śluzowo rzęskowy
-fagocytoza mikrofagów płuc
Drogi oddechowe człowieka wyściełane są nabłonkiem wielorzędowym. Nabłonek zaopatrzony jest w komórki wytwarzające rzęski, pokryty jest śluzem, wytwarzanym przez komórki kubkowe. Śluz en ma wysoką lepkość( zawiera mucynę), ma właściwości bakteriobójcze, dzięki zawartości lizozymu- enzym rozpuszczający ściany komórkowe bakterii Gram+.
Oba typy komórek tworzą aparat śluzowo rzęskowy. Cząstki zawarte w powietrzu przyklejają się do śluzu, następnie są wymiatane do jamy nosowo gardłowej i wydalane ze śliną lub połykane. Ta metoda skuteczna jest dla aerozolu gruboziarnistego.
Bioaerozol drobnoziarnisty może dostać się do pęcherzyków płucnych. Tam może zostać pochłonięty przez obecne tam makrofagi, mające zdolność do fagocytozy.
Inne metody to
-wyłapywanie większych cząstek przez włoski w otworach nosowych
-odruch kaszlowy
-przez hamujące działanie naturalnej mikroflory błony śluzowej dróg oddechowych(interferencja bakteryjna)
Omów czynniki wpływające na stężenie drobnoustrojów w powietrzu.
- wielkość emisji drobnoustrojów, zależna od źródła emisji
-odległość od źródła emisji
-prędkość wiatru
-przeżywalność drobnoustrojów
-opady atmosferyczne
Wielość emisji i skład gatunkowy emitowanego bioaerozolu zależy od źródeł emisji. Są różne czynniki wpływające na stężenie początkowe powstającego bioder. Np. dla komory napowietrzania biologicznej oczyszczalni ścieków są to m. In: stężenie mikroorganizmów w ściekach i sposób ich napowietrzania.
Stężenie musi być odpowiednio duże, bo w przeciwnym razie nie dojdzie do powstania bioaerozolu. Określa to tzw. próg emisj9 dla ścieków 10^3 w 1 cm3).
Powstały bioaerozol rozprzestrzenia się podobnie jak aerozol niebiologiczny, z tą różnicą, że mikroorg z czasem zamierają. Wiejący wiatr rozrzedza aerozol, stężenie na zawietrznej od źródła emisji zmniejsza się.
Dodatkowo stężenie bioaer. zmniejsza grawitacja(działa głównie na większe cząstki), oraz opad atmosferyczny(niekiedy radykalnie redukuje ilość bioaero.)
Na przeżywalność mikroorganizmów w powietrzu wpływa:
-odporność właściwa dla danego mikroorganizmu
-warunki meteorologiczne(wilgotność, temp, promieniowanie, zanieczyszczenia powietrza)
-zanieczyszczenie powietrza
-czas przebywania w powietrzu
-zdolność wytwarzania form przetrwanych
Mokroorganizmy chorobotwórcze przenoszone drogą powietrzną i wywoływane przez nie choroby.
W powietrzu obecne są: bakterie, grzyby, wirusy, (też ich formy przetrwale bakterii- endospory, zarodniki grzybów)
Grupy zagrożeń:
-choroby zakaźne- wirusy, bakterie grzyby, pierwotniaki
-choroby alergiczne
-zatrucia(endotoksyny, mikotoksyny)
Choroby wirusowe
-Grypa-o rtomyxowirusy
-odra, świnka- paramyxowirusy
-zapalenie opon mózgowych- enterowirusy
Choroby bakteryjne:
-gruźlica płuc- prątki gruźlicy Mycobacterium tuberculosis
-zapalenie pluc- gronkowce, pneumokoki Streptococcus pneumoniea
-angina- paciorkowce
-ropne zapalenie dolnych i górnych dróg oddechowych- pleomorficzne pałeczki Haemophilus
-nokardioza- promieniowce z rodzaju Nocardia
-legionellozy- pałeczki z rodzaju Leginella
Choroby grzybowe:
-grzybice powierzchniowe- Microsporum racemosum
-grzybice głęboki: aspergiloza-kropidlak Aspergillus fumigatus; kryptokokoza- drożdżak Cryptococcus neoformans
-astma oskrzelowa- cladosporium
Choroby pierwotniacze:
Nie dotyczą człowieka; np. pryszczyca u zwierząt parzystokopytnych
Dodatkowe informacje:
Grzyby występujące w powietrzu to min: Penicillum, Aspergillus( niger, flavus), Rhizopus microsporus.
Ich obecność w powietrzu może powodować: wady wrodzone, niska masa ciała urodzeniowa, poronienia, bóle głowy, zaburzenia czynności wątroby, dolegliwości neurologiczne, zmiany skórne, podrażnienia błon śluzowych, układu oddechowego oraz nowotwory min: płuc , wątroby. Niektóre grzyby pleśniowe wytwarzają toksyczne metabolity tzw mykotoksyny. Aspergillus flavus wytwarza alfatoksyny, Aspergillus ochraceus - ochratoksyny. Powodują one mykotoksykozy i śmierć.
Bakterie saprofityczne:
-Pseudomonas aeruginosa - gram ujemna, pałeczka, żyje głównie w glebie i wodzie; pałeczka ropy błękitnej Pseudomonas aeruginosa to bakteria, która często atakuje ludzi przewlekle chorych i długo leżących w szpitalu. Jej obecność w płucach wywołuje przewlekły proces zapalny, który w końcu prowadzi do zniszczenia dużych obszarów tkanki płucnej, skurczów oskrzeli wywołanych reakcją zapalną i poważnych kłopotów z oddychaniem.
-Staphylococcus ureus- to gronkowiec złocisty, gram dodatnia bakteria. Charakterystycznymi objawami zatrucia gronkowcem są: wymioty, biegunka, spadek ciśnienia krwi, zapaść a nawet śmierć. Bakteria ta jst odporna na wysokie temperatury. Wywołuje tez zakażenia ropne skóry, czyraki.
Termofilne promieniowce są nitkowatymi, zarodnikującymi bakteriami, które rozwijają się w wysokiej temperaturze, w zanieczyszczonych urządzeniach klimatyzacyjnych. Liczne gatunki mikroorganizmópuje proces samo zagrzewania do temperatury. Np:, Thermoactinomyces thalpophilus , Saccharomonospora viridis , Thermomonospora Fusa, są znaną przyczyną alergicznego zapalenia pęcherzyków płucnych.
Omów bioaerozol jako czynnik alergiogenny.
Wielkość cząstek bioaerozolu a choroby alergiczne:
cząstki powyżej 10ၭm, zatrzymywane w jamie nosowo-gardłowej, powodują katar sienny (np. zarodniki grzyba Alternaria, pyłki traw)
cząstki o średnicy 4-10ၭm, zatrzymywane w oskrzelach, powodują astmę oskrzelową (np. zarodniki grzyba Cladosporium)
cząstki poniżej 4ၭm, przenikające do oskrzelików i pęcherzyków płucnych, wywołują alergiczne zapalenie pęcherzyków płucnych, a także astmę oskrzelową (zarodniki grzybów Aspergillus i Penicillium, większość bakterii, w tym termofilne promieniowce).
Omów toksyny występujące w powietrzu i sposób ich wykrywania.
Endotoksyny
toksyny występujące w błonie zewnętrznej bakterii Gram-ujemnych. Jest to kompleks lipopolisacharydowy wchodzący w skład zewnętrznych warstw komórki bakterii Gram-ujemnych. Endotoksyna jest uwalniana po rozpadzie (lizie) komórki. Na organizm człowieka działają toksycznie, są jednak mniej groźne od egzotoksyn. Wywołują:
- głębokie zaburzenia naczynioruchowe
- gorączkę
- zaburzenia metabolizmu cukrów, tłuszczów i białek
- zaburzenia krzepnięcia
- drażnią skórę
- obniżają fagocytozę
Są stosunkowo trwałe chemicznie, odporne na ogrzewanie w temp. 60 °C przez kilka godzin. Lps jest antygenem i wywołuje odpowiedź immunologiczną.
Wykrywanie toksyn i endotoksyn w powietrzu:
Wykrywanie toksyn i alergenów występujących w powietrzu wymaga często żmudnych badań. Badanie na ich obecność opiera się przede wszystkim na wywoływaniu reakcji immunologicznej z użyciem przeciwciał skierowanych przeciwko znanym antygenom oraz badaniach chromatografi cznych, np. w przypadku mikotoksyn. Wykrywanie endotoksyn występujących w powietrzu wymaga podjęcia następujących działań [2]: przefi ltrowania powietrza przez filtr membranowy (z włókna szklanego lub PCV), rozcieńczania odfi ltrowanych komórek z preparatem z krwi skrzypłocza (morski stawonóg) z dodatkiem substancji chromogennej, wykonania pomiaru wytworzonej lu-minescencji.
Wymień bytowe źródła bioaerozolu i omów jedno z nich.
Z higienicznego punktu widzenia, ważniejsze od naturalnych są bytowe źródła bioaerozolu, związane z działalnością człowieka. Emisje z tych źródeł są niebezpieczne z dwóch powodów:
- mogą rozprzestrzeniać drobnoustroje patogenne (chorobotwórcze)
- często powodują silne zwiększenie liczebności mikroorganizmów w powietrzu, znacznie przekraczające naturalne tło.
Źródła emisji aerozolu biologicznego mogą mieć charakter punktowy (np. komora napowietrzania ścieków) lub powierzchniowy (np. pole nawadniane ściekami).
Do najważniejszych źródeł bioaerozolu należą:
- rolnictwo i przemysł rolno-spożywczy
- oczyszczanie ścieków
- gospodarka odpadami
Rolnictwo i przemysł rolno-spożywczy
Pod względem wielkości emisji jest to najpoważniejsze źródło bioaerozolu, będące efektem intensyfikacji metod produkcji rolnej. Aerozol powstaje podczas wykonywania większości prac rolniczych, np. zbioru plonów, w czasie transportu, przechowywania i przerobu surowców roślinnych i zwierzęcych, oraz w pomieszczeniach hodowlanych.
Zagrożenie dla zdrowia człowieka stwarza tu olbrzymia ilość mikroorganizmów, produktów ich rozkładu i pyłu organicznego, które działają głównie alergicznie i toksycznie. Obecność drobnoustrojów zakaźnych ma tu mniejsze znaczenie.
Do najważniejszych składników tego aerozolu należą:
- grzyby pleśniowe, m.in. tzw. grzyby przechowalniane i tzw. grzyby polowe,
- pałeczki gramujemne, głównie z rodzaju Ervinia,
- promieniowce termofilne,
- pył pochodzenia biologicznego.
Ekspozycja na taki aerozol często powoduje przewlekłe choroby układu oddechowego, np. alergiczne zapalenie pęcherzyków płucnych. Uważa się, że szczególne zagrożenie dla zdrowia powstaje, gdy ponad 50% aerozolu należy do frakcji respirabilnej, a stężenie bioaerozolu przekracza 10^5 cfu/m3. Wielkość ta jest często przekraczana ponad stukrotnie(np. w chlewniach, spichrzach zbożowych).
Wymień i omów mikroorganizmy wskaźnikowe sanitarnej analizy powietrza.
Do stosowanych w sanitarnej analizie powietrza mikroorganizmów wskaźnikowych należą m. in.:
gronkowce hemolizujące,
gronkowce mannitolododatnie i mannitoloujemne,
promieniowce,
pałeczki Pseudomonas fluorescens.
Gronkowce to jedne z najpospolitszych bakterii w przyrodzie. Nie wszystkie są chorobotwórcze, wiele z nich występuje na skórze i błonie śluzowej człowieka nie powodując chorób. Gatunki chorobotwórcze wykazują wysoką aktywność metaboliczną, dzięki której można je odróżnić od niechorobotwórczych.
Gronkowce chorobotwórcze wywołują m. in.:
całkowitą hemolizę krwinek czerwonych (erytrocytów) na agarze z krwią - hemoliza polega na rozkładzie erytrocytów przez pewne toksyny produkowane przez bakterie, co przejawia się powstawaniem charakterystycznych przejaśnień wokół kolonii.
kwaśną fermentację mannitolu na podłożu Chapmana - podłoże Chapmana zawiera 10% NaCl, co powoduje, że wyrastają na nim głównie gronkowce, odporne na duże stężenie soli. Obecność w podłożu mannitolu (wielowodorotlenowego alkoholu) pozwala zróżnicować gronkowce na mannitolododatnie (zdolne do fermentacji mannitolu) i mannitoloujemne (niezdolne). Stwierdzenie hemolizy i fermentacji mannitolu zwiększa prawdopodobieństwo, że wykryte gronkowce są chorobotwórcze.
Promieniowce to typowe bakterie glebowe. W przeciwieństwie do innych bakterii, komórki promieniowców mają często postać rozgałęziających się nitek i mogą tworzyć tzw. pseudogrzybnię, podobnie jak strzępki grzybów. Z tego powodu wytwarzają one charakterystyczne meszkowate kolonie na podłożu Pochona. Wiele promieniowców ma zdolność wytwarzania antybiotyków (promieniowcowym antybiotykiem jest np. streptomycyna). Obecność promieniowców w powietrzu może wskazywać na środowisko glebowe jako źródło zanieczyszczenia powietrza.
Bakteria Pseudomonas fluorescens jest pospolitą bakterią wodną. Na podłożu Kinga B wytwarza ona barwniki fluoryzujące powodujące świecenie kolonii w świetle UV. Obecność tych bakterii w powietrzu może wskazywać na środowisko wodne jako źródło zanieczyszczenia.
Poza tym w badaniu zanieczyszczenia mikrobiologicznego powietrza wokół określonego emitora, wykorzystuje się gatunki charakterystyczne dla tego emitora. Pozwala to na wyznaczenie strefy jego oddziaływania na stan sanitarny powietrza - granicę oddziaływania będzie wyznaczał obszar występowania mikroorganizmu wskaźnikowego. I tak np.:
dla rolnictwa i przemysłu spożywczego:
grzyby pleśniowe, m. in. tzw. grzyby przechowalniane (Aspergillus, Penicillium) i tzw. grzyby polowe (Cladosporium i Alternaria), najpospolitsze grzyby w powietrzu,
pałeczki gramujemne, głównie z rodzaju Ervinia,
promieniowce termofilne,
pył pochodzenia biologicznego (m. in. fragmenty naskórka, pierza, cząstki wydalin i pył roślinny).
dla oczyszczalni ścieków za najbardziej specyficzne uważa się bakterie jelitowe (pałeczki z grupy coli - to przede wszystkim szczepy Escherichia coli oraz drobnoustroje z rodzaju Enterobacter, Citrobacter i Klebsiella, wykrywane są one na podłożach z laktozą po inkubacji w temperaturze 37oC, bakterie grupy coli typu kałowego - termo tolerancyjne - to głównie szczepy Escherichia coli i tylko te nieliczne szczepy z rodzajów Enterobacter, Citrobacter i Klebsiella, które mają zdolność fermentacji laktozy w temperaturze 44 oC; pałeczki durowe) i wirusy jelitowe, gdyż zazwyczaj nie spotyka się ich w powietrzu po stronie nawietrznej oczyszczalni. Z tego powodu uważa się je za mikroorganizmy wskaźnikowe, pomocne w wyznaczaniu strefy oddziaływania oczyszczalni na środowisko.
dla kompostowni - dobrym wskaźnikiem oddziaływania na otoczenie jest pospolity w kompoście grzyb pleśniowy - kropidlak popielaty (Aspergillus fumigatus).
dla składowisk odpadów - dobrymi mikroorganizmami wskaźnikowymi są grzyby ciepłolubne (rosnące w temp. 37oC) i keratynolityczne (rozkładające keratynę).
Na czym polega mikrobiologiczna ocena stanu sanitarnego powietrza?
Ocena stanu sanitarnego powietrza obejmuje aspekt ilościowy i jakościowy, i zależy od rodzaju ocenianego powietrza. Inne kryteria stosuje się do powietrza atmosferycznego, a inne do powietrza wewnątrz różnego typu pomieszczeń. Wartości bezpiecznych stężeń podawane przez różnych autorów różnią się.
Według norm przyjętych w Polsce powietrze atmosferyczne jest czyste, jeśli zagęszczenie komórek bakterii mezofilnych nie przekracza 1000 jtk/m3, a grzybów mikroskopowych 3000 jtk/m3.
Wewnątrz pomieszczeń mieszkalnych ogólna liczba bakterii mezofilnych nie powinna przekraczać 2000 jtk/m3, a grzybów 300 jtk/m3.
Jeśli zagęszczenie mikroorganizmów przekracza podane wartości, lub w skład aerozolu wchodzą drobnoustroje niebezpieczne dla człowieka, to takie powietrze uważa się za zanieczyszczone mikrobiologicznie.
Dla pomieszczeń użytkowych dopuszczalne wartości graniczne zależą od ich przeznaczenia, np. na sali operacyjnej nie może być w ogóle grzybów a liczba bakterii nie może przekraczać 100 cfu/m3. Natomiast np. w chlewni dopuszcza się stężenie bakterii wynoszące 200000 cfu/m3, a grzybów 10000 cfu/m3.
Bardzo ważna z higienicznego punktu widzenia jest znajomość wielkości frakcji respirabilnej bioaerozolu. Im większy jest jej udział procentowy, tym większe zagrożenie dla zdrowia stwarza powietrze, nawet jeśli nie ma w nim mikroorganizmów wywołujących choroby zakaźne. Wdychanie takiego powietrza może bowiem być powodem alergii, zatruć i pylic.
Badania jakościowe z konieczności muszą być ograniczone do mikroorganizmów wskaźnikowych, bowiem identyfikacja drobnoustrojów chorobotwórczych jest zwykle żmudna i droga. Gatunki wskaźnikowe same nie muszą być chorobotwórcze, ale ich występowanie wskazuje pośrednio na potencjalne zagrożenie mikroorganizmami patogennymi.
Na czym polega biofiltracja gazów i kiedy może być stosowana?
Biologiczne oczyszczanie powietrza w biofiltrach polega na powolnym przepuszczaniu gazów przez warstwę materiału porowatego zasiedlonego przez mikroorganizmy. W określonych warunkach pracy biofiltra, zanieczyszczenia obecne w gazie wylotowym są absorbowane i ulegają stopniowemu rozkładowi na naturalne substancje takie jak woda i dwutlenek węgla. Początkowo zanieczyszczone powietrze jest poddane wstępnemu oczyszczaniu w zintegrowanym z biofiltrem wstępnym skruberze, gdzie zostają zapewnienie optymalne warunki niezbędne dla dalszego procesu oczyszczania powietrza. W wstępnym skruberze zanieczyszczony gaz zostaje ochłodzony do odpowiedniej temperatury, odpowiednio nawilżony oraz pozbawiony stałych cząsteczek. Wstępny skruber pełni również rolę buforudla pojawiających się w powietrzu wysokich stężeń zanieczyszczeń. Wstępnie przygotowane powietrze rozprowadzane jest w kanale dystrybucyjnym a następnie przepływa z małą prędkością przez biologiczne złoże organiczne. Jako materiał filtrujący najczęściej stosuje się mieszaniny surowców pochodzenia organicznego, zawierające duży ładunek biomasy. Sposób ułożenia materiału filtrującego powinien zapewniać jego równomierne napowietrzenie i gwarantować kontakt całego strumienia gazu ze złożem. W celu zapewnienia odpowiednich warunków pracy biofiltra jest konieczne, aby materiał organiczny posiadał jednolitą strukturę oraz wystarczającą wilgotność. Zastosowanie biofiltrów jako efektywnego procesu oczyszczania powietrza ma miejsce głownie w miejskich i przemysłowych oczyszczalniach ścieków, instalacjach suszenia osadów, przemyśle spożywczym i produkcyjnym żywności, cukrowniach, rzeźniach, przemyśle przetwórstwa mięsnego i rybnego, fermach hodowlanych, kompostowniach i wysypiskach śmieci, przemyśle chemicznym, lakierniach oraz wielu innych procesach technologicznych. Wiele związków organicznych takich jak fenole, formaldehyd, ksylen, toluen, styren, alkohole, ketony i glikole ulegają efektywnemu rozkładowi. Przy zachowaniu optymalnych parametrów pracy likwidacja zanieczyszczeń jest bardzo wysoka i wynosi 98%.
Podaj kryteria gatunku i omów zasady nazewnictwa prokariotów.
Kuliste:
-ziarniaki(Coccus)
-dwoinki(Diplococcus)
-paciorkowce(Strepotococcus)
-gronkowce(Staphylococcus)
-pakietowce(Sarcina)
Wydłużone, cylindryczne:
-laseczki(Bacillus)
-pałeczki(Bacterium)
-maczugowce(Corynebacterium)
Skręcone spiralnie:
-przecinkowce(Vibrio)
-śrubowce(Spirillum)
-krętki(Spirochaeta)
Rozgałęzione:
-prątki(Mycobacterium)
-promieniowce(Actinomyces)
Kryteria gatunku : reakcja Gram (dodatnie lub ujemne), tolerancja na temperaturę (mezofile i psychrofile), morfologia

Porównaj 3 domeny organizmów.
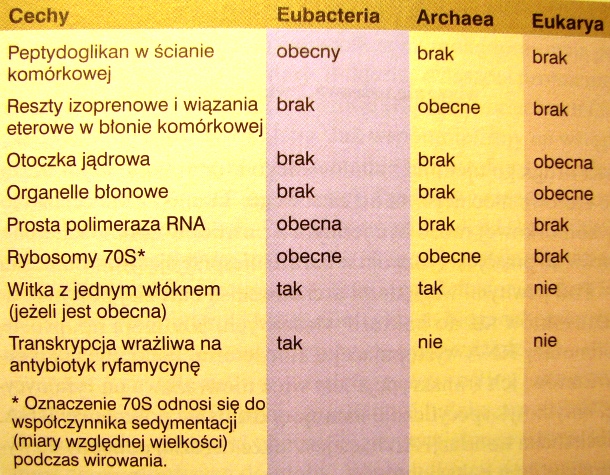
Na czym polega taksonomia naturalna i jakie grupy bakterii wyróżnia się na jej podstawie?
Taksonomia naturalna (filogenetyczna)-połączenie spokrewnionych ze sobą form mających wspólnych przodków i uporządkowanie ich w jedno wspólne drzewo filogenetycznie bakterii. Te same właściwości biochemiczne (sekwencje aminokwasów w enzymach spełniających podobne funkcje)odznaczają się konserwatywną budową taką jak rybosomowe kwasy nukleinowe.
FILOGENETYCZNE GRUPY BAKTERII
Na podstawie sekwencji 16S rRNA wyróżnia się 12 głównych linii bakterii:
• Aguifex - Hydrogenobacter: hipertermofilne chemolitotrofy utleniające H
2 lub redukujące związki siarki, żyją w temperaturze do 95°C
• Thermotoga: hipertermofile, większość gatunków jest chemoorganotroficzna, z reguły o
beztlenowym metabolizmie fermentacyjnym
• Zielone bakterie niesiarkowe (grupa Chloroflexus) - większość to fototrofy.
• Deinococcus (wiele gatunków wyjątkowo odpornych na promieniowanie) oraz
termofile, przedstawiciel: Thermus aquaticus
• Krętki (spirochetes)
• Zielone bakterie siarkowe (fototrofy)
• Bacteroides
- Flavobakterie:
mieszane typy fizjologiczne od bezwzględnych
beztlenowców (Bacteroides) do bezwzględnych tlenowców (Sporocytophaga) przez
względnie tlenowe.
• Grupa Planctomyces - Pirella; reprodukcja przez pączkowanie, brak peptydoglikanu w
ścianie komórkowej.
• Chlamydie i Mykoplazmy:
brak
ściany
komórkowej,
obligatoryjne,
wewnątrzkomórkowe pasożyty komórek zwierzęcych.
• Bakterie Gram-dodatnie
- o niskiej zawartości GC
- o wysokiej zawartości GC
• Sinice
• Bakterie purpurowe (Protobakterie): największa i najbardziej zróżnicowana grupa
prokariontów
Na czym polega taksonomia sztuczna i numeryczna?
Taksonomia sztuczna -jej celem jest grupowanie organizmów zgodnie z ich podobieństwem, tak aby mogłabyć klasyfikowana i oznaczona. W tym systemie posługujemy się kluczem który zawiera nazwy,opisy cech morfologicznych, fizjologicznych.
Taksonomia numeryczna -systematyczne uszeregowanie bakterii. Opiera się na zasadzie Adansona wg którego wszystkie uchwytne cechy organizmu są w klasyfikacji jednakowo ważne. Należy brać pod uwagę możliwie jak najwięcej cech, które ocenia się na + lub -.Oceny odpowiednich kombinacji cech można dokonać za pomocą komputera,każdą cechę danego szczepu porównuje się z tą samą cechą wszystkich pozostałych szczepów.
Omów replikację DNA u mikroorganizmów prokariotycznych i eukariotycznych.
Replikacja DNA to proces, w którym podwójna nić DNA ulega skopiowaniu. Replikacja jest semikonserwatywna - w każdej z dwóch uzyskanych podwójnych nici DNA będzie jedna nić macierzysta i jedna nowa.
Substratami tego procesu są: DNA i trifosforany nukleozydów. W procesie tym bierze udział wiele enzymów, w tym: helikaza - tnie wiązania wodorowe między niciami DNA umożliwiając rozpoczęcie kopiowania primaza - syntetyzuje primer (starter) - krótki fragment z RNA umożliwiający rozpoczęcie procesu polimeraza DNA - z trifosforanów nukleozydów syntetyzuje (dobudowuje na zasadzie komplementacji) brakujące nici DNA egzonukleaza - usuwa primery RNA z nici, bez niej nowa nić zawierałaby zarówno DNA jak i niewielkie ilości RNA.
ligaza DNA - uzupełnia brakujące wiązania fosfodiestrowe w szkielecie nowo-zsyntezowanej nici DNA różne enzymy pomocnicze
Zasady replikacji są podobne u wszystkich organizmów, przy czym największe różnice występują między bakteriami z jednej strony, a prokariotami i eukariontami z drugiej.
U bakterii replikacja zaczyna się w ustalonym miejscu i postępuje bardzo szybko, z prędkością rzędu 1000 nukleotydów na sekundę.
U eukariotów replikacja jest o wiele wolniejsza, ok. 50 nukleotydów na sekundę, jednak zachodzi równocześnie w wielu miejscach. Polimeraza DNA działa jedynie w kierunku od końca 3' do końca 5'. Z tego powodu jedna z nici jest syntezowana w sposób ciągły, druga (ta, którą chcielibyśmy zsyntezować w przeciwną stronę) fragmentami.
Przebieg replikacji DNA
W punktach
1) Dwie nici podwójnej helisy w pewnym miejscu rozplatają się; powoduje to rozerwanie par zasad komplementarnych
W pobliżu miejsca rozplecenia zawsze znajduje się dostateczna ilość wolnych nukleotydów; do każdej z zasad w rozplecionych niciach dołącza zasada komplementarna, stanowiąca część wolnego nukleotydu - jednostki budulcowej DNA
Między nowo dołączonymi nukleotydami powstają trwałe połączenia - w ten sposób tworzy się nowa nić DNA.
Szczegóły procesu
Kopiowanie podwójnej helisy DNA jest procesem złożonym. Rozpoczyna się w miejscu inicjacji, liczącym ok. 200-300 par nukleotydów. Aby replikacja przebiegła prawidłowo, rozdzielenie obu nici musi zajść bez zniszczenia ich struktury podstawowej (I-rzędowej) i w odpowiednich warunkach, jak: dokładne odczytanie matrycy DNA, obecność odpowiedniej liczby wolnych nukleotydów, zachowania komplementarności. Na koniec musi dojść do terminacji replikacji, ewentualnego uzupełnienia braków na końcu nowopowstałej cząsteczki i połączenia nowych cząsteczek w helisę.
U bakterii zakończenie replikacji jest niemal automatyczne (po skopiowaniu całego kolistego DNA , który jest pojedynczym replikonem.
U eukariotów miejsc replikacji (replikonów) jest wiele, a jej terminacja zachodzi po zakończeniu wielu procesów replikacyjnych zachodzących niemal jednocześnie w różnych miejscach replikujących cząsteczek DNA. Do terminacji dochodzi, gdy widełki replikacyjne replikonu natkną się na specjalną sekwencję terminacyjną. Proces replikacji nie-kolistych (eukariotycznych) cząsteczek DNA wiąże się z problemem wolnych zakończeń powstających cząsteczek DNA. Zakończenia te, zwane telomerami składają sie z krótkich, ale wielokrotnie powtórzonych sekwencji. Replikazy wydłużają jedynie istniejące już nici, nie są natomiast w stanie zsyntetyzować końcowych odcinków telomerów. W rezultacie odcinki te narażone są na regularne skracanie. Skracaniu temu zapobiega obecność telomerazy, która przeprowadza odwrotną transkrypcję tych odcinków, posługując się jako "matrycą" nie DNA, ale RNA, będącym częścią składową tego enzymu. Zapobiega to odsłonięciu znaczących fragmentów DNA.
Różnice między grupami organizmów
Między replikacją u bakterii oraz prokariotów i eukariotów (które prawie zawsze mają wspólne mechanizmy przetwarzania informacji) zachodzą dość duże różnice. Nie wszystkie enzymy uczestniczące w replikacji DNA są uważane za homologiczne, co może sugerować, że ostatni uniwersalny wspólny przodek miał jedynie częściowo wykształcony proces replikacji DNA.
Omów transkrypcję u mikroorganizmów eukariotycznych i prokariotycznych.
Omów translację u mikroorganizmów eukariotycznych i prokariotycznych.
Wymień znane rodzaje mutacji i omów sposób ich powstawania.
Wyjaśnij mutagenne działanie promieniowania UV.
Światło ultrafioletowe działa na mikroorganizmy zarówno letalnie, jak i mutagennie. Mechanizm tego działania nie został jeszcze w pełni poznany. Znaczna współzależność między widmem absorbowanym przez kwasy nukleinowe a widmem mającym efekt letalny i mutagenny sugeruje, że to właśnie kwas nukleinowy jest celem ataku przez UV. Blisko ultrafiolet, około 260 nm, jest najbardziej aktywny. Toksyczne działania uboczne prawie nie występują. UV niszczy głównie zasady pirymidynowe. Dwie tyminy sąsiadujące ze sobą zostają połączone wiązaniami kowalencyjnymi. Takie dimery tyminowe mogą następnie powodować błędy repikacyjne. Fotodimery pirymidyn powodują zahamowanie włączania nukleotydów w procesie replikacji prze polimerazę DNA III co powoduje fragmentację DNA i efekt letalny.
Odkrycie zdolności do naprawy przynajmniej części uszkodzeń DNA jest związane z działaniem promieni UV na bakterie. Po naświetleniu zawiesiny bakteryjnej dużą dawką UV i następnie inkubacji w ciemności, tylko mały odsetek komórek zachowuje zdolność do tworzenia kolonii. Przeciwnie, jeżeli bakterie po naświetlaniu UV poddamy działaniu światła o dłuższych falach, procent przeżywających komórek wzrasta o kilka rzędów wielkości. Ta fotoreaktywacja zachodzi z udziałem enzymu fotoliazy, który czerpiąc energię ze światła o dłuższych falach, rozszczepia dimery tyminy na monomery. W ten sposób następuje powrót do stanu wyjściowego, a naprawa jest bezbłędna.
Drugi, równie bezbłędny sposób naprawy uszkodzeń powodowanych promieniowaniem nie wymaga światła. Ta ciemna reaktywacja polega na wycięciu uszkodzonego kawałka nici DNA i zastąpieniu go nowymi nukleotydami.
Skuteczność naprawy uszkodzeń radiacyjnych zależy od szczepu bakterii.
Wyjaśnij zjawisko rewersji mutacji.
Mutacje powrotne albo rewersje: Wiele mutantów, choć nie wszystkie, może mutować wstecznie, z przywróceniem cech dzikiego rodzica. Odróżnia się dwa rodzaje mutacji pierwotnych:
1)rewertanty (prawdziwe), gdy druga mutacja w tym samym genie odtwarza genotyp pierwotny, tj. genotyp typu dzikiego istniejący zanim zaszła pierwsza z mutacji
2)rewertanty funkcjonalne z zachowaną oryginalną mutacją, lecz fenotypowo dzikie w wyniku powstania drugiej mutacji w innym locus (określony obszar chromosomu zajmowany przez gen) w obrębie tego samego lub nawet innego genu. Do grupy tej należą mutanty supresorowe, w których nowa mutacja hamuje w sposób pośredni początkowy defekt. Mutanty supresorowe niejednokrotnie rosną wolniej niż prawdziwy typ dziki. Podstawą wolniejszego wzrostu jest zachodzące od czasu do czasu włączanie złego aminokwasu wskutek możliwości wykorzystania kilku pokrewnych tRNA przez jeden aminokwas.
Omów proces rekombinacji.
Rekombinacja jest to proces wymiany materiału genetycznego, w wyniku którego powstają nowe genotypy. Zgodnie z obecnym stanem wiedzy, istnieją dwa różne mechanizmy, za pomocą których obcy DNA pobrany przez komórkę bakteryjną może zostać włączony do chromosomu lub plazmidu, są to:
rekombinacja ogólna lub homologiczna
rekombinacja zależna od miejsca lub sekwencji, zwana również rekombinacją zlokalizowaną
-Rekombinacja ogólna lub homologiczna-terminem tym określamy proces, w którym obcy DNA znajdujący się w komórce zostaje włączony do DNA gospodarza przez połączenie się w pary homologicznych sekwencji, pęknięcie i krzyżową wymianę (crossing-over). Wzajemna wymiana odcinków DNA między chromosomami dawcy i biorcy rozpoczyna się równoległym ułożeniem (sparowaniem) homologicznych odcinków i jednoniciowym pęknięciem. Następnym etapem jest wymiana jednoniciowych segmentów DNA dawcy i biorcy. W procesie tym bierze udział białko RecA i białko wiążące jednoniciowy DNA. Efekt rekombinacji ujawnia się dopiero w komórkach potomnych. Warunkiem koniecznym tego procesu jest istnienie długiego obszaru homologii sekwencji w DNA dwóch partnerów. Wiadomo jednak, że niewielkie różnice sekwencji, na przykład powstałe w wyniku mutacji, bywają tolerowane. Homologiczna rekombinacja może więc zachodzić między DNA dzikiego typu i DNA mutanta. Rekombinacje ogólną katalizuje co najmniej 6 enzymów mających również znaczenie w reparacji DNA. Najważniejszym z nich jest wspomniany wcześniej RecA.
-Rekombinacja zależna od określonego miejsca lub sekwencji-ten rodzaj rekombinacji, w przeciwieństwie do rekombinacji ogólnej, wymaga jedynie krótkiego odcinka homologii DNA koniecznej do rozpoznania się partnerów. Rekombinacja ta nie jest katalizowana przez białko RecA, lecz przez enzymy swoiste dla rekombinujących cząsteczek DNA. Jeśli sekwencja wymagana do rozpoznania znajduje się w dwóch partnerach mamy do czynienia z „rekombinacją określoną przez dwa miejsca”. Przykładem tego typu rekombinacji jest integracja (włączenie) bakteriofaga lambda i plazmidu płciowego F do chromosomu E.coli. Obecność rozpoznawanej sekwencji tylko w jednej cząsteczce DNA prowadzi do rekombinacji „określonej przez jedno miejsce”, która jest związana z ruchomymi elementami genetycznymi, jak sekwencje insercyjne, transpozony, bakteriofag Mu.
Jak wykazały doświadczenia genetyczne, przejście faga lamba w stadium profaga jest związane z jego integracją w określone miejsce chromosomu E.coli, między operonami gal i bio. Kolisty chromosom faga wiąże się przy pomocy białek w określonym miejscu attP do miejsca attB chromosomu gospodarza znajdującego się między genami bio i gal. Następnie chromosom faga zostaje zintegrowany przez pęknięcie i ponowne wzajemne połączenie dwóch nici. Prowadzi to do integracji fagowego DNA z DNA gospodarza.
Co to są plazmidy i jakie pełnią funkcje?
Plazmid - cząsteczka DNA występująca w komórce poza chromosomem i zdolna do autonomicznej (niezależnej) replikacji. Plazmidy występują przede wszystkim u prokariotów, ale znane są także nieliczne plazmidy występujące u eukariotów. Zazwyczaj plazmidy nie niosą genów metabolizmu podstawowego, a więc nie są komórce niezbędne do przeżycia. Mogą jednak kodować produkty potrzebne w pewnych specyficznych warunkach, na przykład geny oporności na antybiotyki lub umożliwiające rozkład i asymilację różnych związków odżywczych. Plazmidy w drodze koniugacji mogą być przekazywane pomiędzy komórkami bakteryjnymi.
Omów mechanizmy przenoszenia materiału genetycznego u bakterii.
Bakterie posiadają zwykle pojedynczy chromosom, którego wielkość wynosi zaledwie od około 16 000 par zasad u endosymbiotycznych Candidatus Carsonella ruddii[78] do 12 000 000 par u Sorangium cellulosum[79]. Krętki z rodzaju Borrelia są jedynym wyjątkiem, gdyż mają pojedynczy chromosom o kształcie lini. Należy tu Borrelia burgdorferi, który wywołuje boreliozę, zwaną także krętkowicą kleszczową oraz chorobą z Lyme[80]. Bakteryjne geny zwykle są umieszczone na pojedynczym odcinku DNA i chociaż u bakterii zdarzają się także różne typy intronów, to są one o wiele rzadsze niż u eukariontów[81]. Bakterie mogą także posiadać plazmidy, które są dodatkowymi, małymi chromosomami. Zawierają one DNA odpowiadające między innymi za oporność na antybiotyki, oraz czynniki decydujące o wirulencji patogenu.
Bakterie potrafią także wymieniać się genami między sobą. Zjawisko to może nastąpić na trzy różne sposoby. Po pierwsze bakterie mogą pobrać dany gen z środowiska w procesie nazywanym transformacją. Drugą metodą jest przeniesienie genów w procesie transdukcji, kiedy bakteriofag wprowadza swoje geny do chromosomu. Trzecią możliwość stanowi koniugacja bakteryjna, gdzie DNA jest przenoszone poprzez bezpośredni kontakt komórek bakterii.
Jednym z typów bakteryjnego genomu jest połączenie genów bakterii z materiałem genetycznym bakteriofagów. Wiele bakterii jest atakowanych przez te wirusy, które "wstrzykują" do ich chromosomów własny materiał genetyczny. Bakteriofagowe geny mają wielki wpływ na fenotyp bakterii. Dzięki temu na drodze ewolucji niegroźne bakterie zamieniały się w niebezpieczne dla życia innych organizmów, takie jak Escherichia coli O157:H7 czy Clostridium botulinum po przejęciu niesionych przez faga genów pewnej toksyny[82]. Bakterie potrafią jednak bronić się przed zainfekowaniem przez fagi. Służy im do tego specjalny system obronny nazywany systemem ograniczenia modyfikacji, który potrafi degradować obce DNA i rozbijać je na pojedyncze nukleotydy[83]. System ten używa także odpowiednich kolejności CRISPR, dzięki któremu może zapamiętywać szczególne cechy danego bakteriofaga i w późniejszym kontakcie z nim znacznie lepiej go zwalczać, poprzez blokowanie syntezy materiału genetycznego faga i bakterii oraz dalszemu uniemożliwieniu przeprowadzenia intenferencji RNA[84][85]. CRISPR stanowi więc odporność nabytą bakterii na infekcje wirusową.
Bakterie jako organizmy rozmnażające się bezpłciowo dziedziczą identyczny materiał genetyczny od ich rodziców (można nawet powiedzieć, że są ich klonami). Mimo to każda bakteria kształtuje swój własny fenotyp, co jest spowodowane zmianami w DNA. Poza tym wśród bakterii można sztucznie spowodować zmianę. Naturalnie może ona powstać na skutek mutacji oraz rekombinacji genetycznej. Mutacje powstają na skutek błędów w czasie tworzenia repliki DNA lub jako skutek oddziaływania mutagenu. Szansa na powstanie i czas potrzebny do zajścia mutacji są różne w obrębie każdego gatunku, a nawet tej samej bakterii[86]. Genetyczne zmiany u bakterii są powodowane mutacjami przy replikacji genów albo na skutek różnorodnych "nacisków" ze strony człowieka, gdy na wybrany gen oddziałuje się licznymi mutagenami, prowadzi to do zakłócenia procesów wewnętrznych i w konsekwencji do mutacji[87].
Podczas koniugacji jedna komórka („dawca”) wytwarza rurkowate cytoplazmatyczne wyrostki, tzw. pilusy, umożliwiające kontakt między komórkami bakterii. Po wymianie cytoplazmy wraz z materiałem genetycznym (plazmidami) komórki rozdzielają się. Proces ten ma różne odmiany. Wszystkie sposoby wymiany materiału genetycznego nazywają się poziomym transferem genów. Bakterie mogą wykorzystywać wszystkie te metody w naturalnym środowisku[88]. Transfer genowy jest szczególnie cenny przy wytwarzaniu oporności na antybiotyki, gdyż umożliwia bakterii odpornej na działanie antybiotyku uodparnianie innych, poprzez przekazywanie im genów warunkujących odporność, przez co może doprowadzić do uodpornienia całej populacji[89]. Z tego powodu poziomy transfer genów może być z medycznego punktu widzenia niezwykle groźny, gdy zachodzi wśród bakterii chorobotwórczych. Odkrycie poziomego transferu genów (transformacji u dwoinki zapalenia płuc) przez Fredericka Griffitha przyczyniło się do rozwoju genetyki molekularnej i później pozwoliło wyjaśnić rolę DNA.
Wyjaśnij działania operonu.
Operon laktozowy Escherichia Coli jako przykład regulacji ekspresji genu u Procaryota
1. Operon jest to grupa kilku genów, które są wspólnie regulowane. Białka kodowane przez geny operonu są enzymami katalizującymi w komórce kilka, następujących po sobie reakcji chemicznych (tzw. ciąg metaboliczny).
Geny operonu leżą tuż obok siebie, dzięki temu komórka może "uruchomić" je wszystkie "za jednym zamachem" kiedy będą potrzebne - tzn. gdy "opłaca się" przeprowadzenie danego ciągu reakcji. Geny operonu, nazywane również genami struktury, są przepisywane na wspólny mRNA, powstaje jeden długi transkrypt, zawierający informacje o kilku białkach. Transkrypt ten ulega zaraz translacji; sekwencje kodujące białka rozdzielone są sygnałami startowymi i końcowymi dla translacji, dlatego w wyniku tego procesu powstaje od razu kilka osobnych białek . Wspólny transkrypt dla kilku genów kontrolujących dany ciąg reakcji - tak w jednym zdaniu można określić istotę operonu.
Operony wykryto dotychczas wyłącznie u organizmów prokariotycznych; taki sposób regulacji ekspresji kilku genów prawdopodobnie w ogóle nie występuje u organizmów eukariotycznych ze względu na inną budowę genów (są długie i zawierają introny) oraz przestrzenny i czasowy rozdział transkrypcji i translacji. Prokariotyczne mRNA ulega translacji jeszcze podczas trwania transkrypcji, natomiast u organizmów eukariotycznych z pierwotnego tanskryptu usuwane są introny, po czym powstały mRNA wędruje przez pory w błonie jądrowej do cytoplazmy. Długi transkrypt zawierający informacje o kilku białkach prawdopodobnie nie miałby szans na przeżycie w komórce eukariotycznej, uległby zniszczeniu jeszcze na terenie jądra komórkowego, podczas wycinania intronów lub podczas wędrówki z jądra do cytoplazmy.
2. Operon laktozowy występuje u bakterii Escherichia coli. Funkcjonowanie tego operonu zostało wyjaśnione w 1961 roku przez dwóch francuskich uczonych, Francoisa Jacoba i Jacuesa Monoda. Operon laktozowy zawiera trzy geny, kodujące białka związane z metabolizmem laktozy. Pierwsze z nich, permeaza, jest odpowiedzialne za transport cząsteczki laktozy do wnętrza komórki bakterii. Kolejne białko, b-galaktozydaza, rozcina cząsteczkę tego dwucukru na glukozę i galaktozę. Trzecie białko, tansacetylaza, uczestniczy również w metabolizmie laktozy. Wszystkie te białka są potrzebne komórce bakteryjnej tylko wtedy, gdy w jej otoczeniu znajduje się laktoza, zaś innych wydajniejszych żródeł energii (np. glukozy) brak.
3. Operon laktozowy jest regulowany na dwa sposoby; negatywnie i pozytywnie. Regulacja negatywna polega na stałym (czyli konstytutywnym) blokowaniu transkrypcji kiedy w otoczeniu komórki bakteryjnej nie ma laktozy. Regulacja negatywna jest zabezpieczeniem przed produkcją enzymów, które nie są potrzebne komórce, gdy nie ma substratu (laktozy).
Regulacja pozytywna polega na aktywowaniu transkrypcji przez związki, które umożliwiają przyłączenie się polimerazy RNA; zachodzi gdy w otoczeniu bakterii pojawia się laktoza, ale brak glukozy. Glukoza jest bardziej ekonomicznym żródłem węgla, niż laktoza, ponieważ laktoza musi być rozcięta na cukry proste, nim wejdzie w proces glikolizy, a to wymaga energii. Dlatego komórce, która ma do wyboru glukozę i laktozę nie opłaca się "zjadać" laktozy - w takiej sytuacji geny operonu laktozowego nie ulegają włączeniu, mimo obecności laktozy.
Regulacja ekspresji genów operonu laktozowego jest możliwa dzięki odcinkom znajdującym się przed sekwencją kodującą.Odcinki te to: promotor, czyli miejsce wiązania polimerazy RNA, katalizującej transkrypcję oraz operator. Operator jest to krótka sekwencja nukleotydów, znajdująca się tuż przed pierwszym genem struktury.
4. Regulacja negatywna operonu laktozowego. Do sekwencji operatora może przyłączyć się tzw. represor, białko, które blokuje transkrypcję genów struktury, ponieważ jest tak duże, że przyłączone do operatora, zasłania promotor i uniemożliwia przyłączenie się do niego polimerazy RNA.
Gen kodujący represor, czyli tzw. gen regulatorowy, nie należy do operonu laktozowego, znajduje się w innej częsci bakteryjnego genomu, daleko - bo aż kilka tysiecy par zasad od operatora. Gen kodujący represor jest genem konstytutywnym, oznacza to, że ulega on stale ekspresji i niewielka ilość represora zawsze znajduje się w komórce. Represor dociera do operatora na zasadzie dyfuzji i łącząc się z nim blokuje ekspresję genów operonu.
Represor operonu laktozowego jest białkiem allosterycznym - to znaczy takim, które może zmieniać swoją strukturę przestrzenną (konformację) pod wpływem przyłączanych czynników regulacyjnych. Takim czynnikiem może być pochodna laktozy - allolaktoza. Jeżeli w otoczeniu komórki bakteryjnej pojawi się laktoza, niektóre jej cząsteczki zostaną przetransportowane do wnętrza komórki i przetworzone na allolaktozę. Allolaktoza przyłącza się do represora i to doprowadza do takiej zmiany konformacji represora, że nie może on rozpoznać sekwencji operatora i specyficznie się z nią połączyć. Represor przestaje blokować transkrypcję genów struktury.
Laktoza jest w tym wypadku induktorem transkrypcji, pod jej wpływem zablokowane przez represor geny struktury ulegają "włączeniu" (indukcji) ich ekspresja wzrasta tysiąckrotnie; poziom b-galaktozydazy , enzymu odpowiedzialnego za rozcinanie laktozy, znikomy przed indukcją, wzrasta do trzech tysięcy cząsteczek w komórce, co stanowi ok.3% białek komórki Escherichia coli.
5. Regulacja pozytywna operonu laktozowego jest niezbędna, by zaszła transkrypcja. W pobliżu promotora operonu laktozowego znajduje się sekwencja nukleotydów, do której przyłącza się białko aktywujące transkrypcję - tzw. białko CAP - białkowy aktywator katabolizmu (spalanie laktozy jest procesem katabolicznym). Bez tego białka polimeraza RNA ma słabe powinowactwo do promotora i transkrypcja nie zachodzi, nawet w obecności laktozy, przy odblokowanym operatorze.
Białko CAP jest dimerem (tzn. składa się z dwóch podjednostek), podobnie jak represor ma zdolność do zmiany konformacji pod wpływem przyłączenia czynników regulacyjnych. Białko CAP przyjmuje aktywną postać po przyłączeniu cAMP - cyklicznego adenozynomonofosforanu i dopiero wtedy może przyłączyć się do DNA( cAMP reguluje aktywność wielu ważnych dla komórki białek, obrazuje to laboratoryjne powiedzonko: "jeżeli wszystko zawodzi, sprawdź poziom cAMP"). Poziom cAMP jest związany z ilością glukozy w komórce - im mniej glukozy, tym więcej cAMP. Tak więc białko CAP jest aktywne przy niskim stężeniu glukozy.
Dzięki takiej precyzyjnej regulacji operon laktozowy jest uruchamiany tylko w obecności laktozy i przy niskim stężeniu glukozy.
Wyjaśnij proces represji katabolicznej.
Represja kataboliczna - polega na zahamowaniu ekspresji genów kodujących białko enzymatyczne, katalizujące rozkład substancji pokarmowych, na skutek wzrostu dostępności innego substratu, którego katabolizm jest bardziej wydajny energetycznie. Przykładem jest zahamowanie genów operonu laktozowego u pałeczki okrężnicy w obecności glukozy w pożywce.
Mechanizm represji katabolicznej opiera się na fakcie, że polimeraza RNA łączy się z promotorem w operonie lac dużo lepiej w obecności specyficznego białka CAP (catabolite gene activator protein), które musi być związane ze specyficznym miejscem DNA położonym w pobliżu tzw.CBS (CAP binding site). Białko CAP wiąże się z tym miejscem jeśli do tego białka przyłączą się cząsteczki cAMP, co zachodzi przy braku glukozy.
Wyszukiwarka
Podobne podstrony:
Mikrobiologia wykład IV cz 1
mikrobiologia cz 1
mikrobiologia cz 3
grzyby cz 1, Mikrobiologia wet, sem 2
Mikrobiologia+wykład+IV+cz 2
Metody stosowane w diagnostyce mikrobiologicznej cz
mikrobiologia cz 4
mikrobiol cz 1
mikrobiologia cz 1
WIRUSY II cz., Mikrobiologia
mikrobiologia cz.1, pwr, W7 wydział inżynierii środowiska, Pwr OŚ Ochrona Środowiska, Semestr 3, Mik
mikrobiologia cz 2
Metody ilościowe w mikrobiologicznych badaniach żywności Cz 2
Metody ilościowe w mikrobiologicznych badaniach żywności Cz 1
więcej podobnych podstron