
Programowana śmierć
i
starzenie
komórek roślinnych

Wstęp i definicje
• Programowana śmierć komórki (PCD)
jest uznawana
za jedno z najważniejszych zjawisk odpowiedzialnych
za formowanie organów roślinnych.
• PCD jest procesem niezbędnym na wielu etapach
ontogenezy
.
• Przykłady PCD:
– embriogeneza – degradacja komórek tworzących tapetum,
bielma i wieszadełka,
– histogeneza – powstawanie
aerenchymy
(miękiszu
powietrznego), powstawanie członów naczyń
ksylemu
(kanałów transportujących wodę w roślinie)
– reakcja nadwrażliwości (HR) – bardzo szybkie obumieranie
komorek zainfekowanych patogenem.

• Programowana śmierć komórki (programmed cell death,
PCD)
- aktywny proces eliminacji składników komórki,
prowadzący ostatecznie do jej śmierci, zachodzący
podczas rozwoju, a także w odpowiedzi na warunki
środowiskowe.
• Starzenie
– podczas starzenia, z komórek organów
które obumierają zostają wycofywane zmagazynowane
zasoby węgla i azotu do pozostałych części rośliny.
• Zasadnicza różnica
–
starzenie
może być procesem
odwracalnym
, jednak po przekroczeniu określonej
granicy staje się
nieodwracalnym procesem PCD.
Wstęp i definicje

Różnica PCD w komórkach zwierzęcych
i roślinnych
• W komórkach zwierzęcych przed PCD nie ma
wycofywania zmagazynowanych zasobów węgla i azotu
do tkanek żywych.
Starzenie komórek zwierzęcych nie musi się wiązać z ich
śmiercią. Stara komórka nie dzieli się, zmienia się jej
morfologia i metabolizm, ale może długo funkcjonować i
żyć.
• W komórkach roślinnych oba procesy są
zsynchronizowane, starzenie uznaje się jako
zaprogramowaną genetycznie, samobójczą
śmierć komórki - PCD.
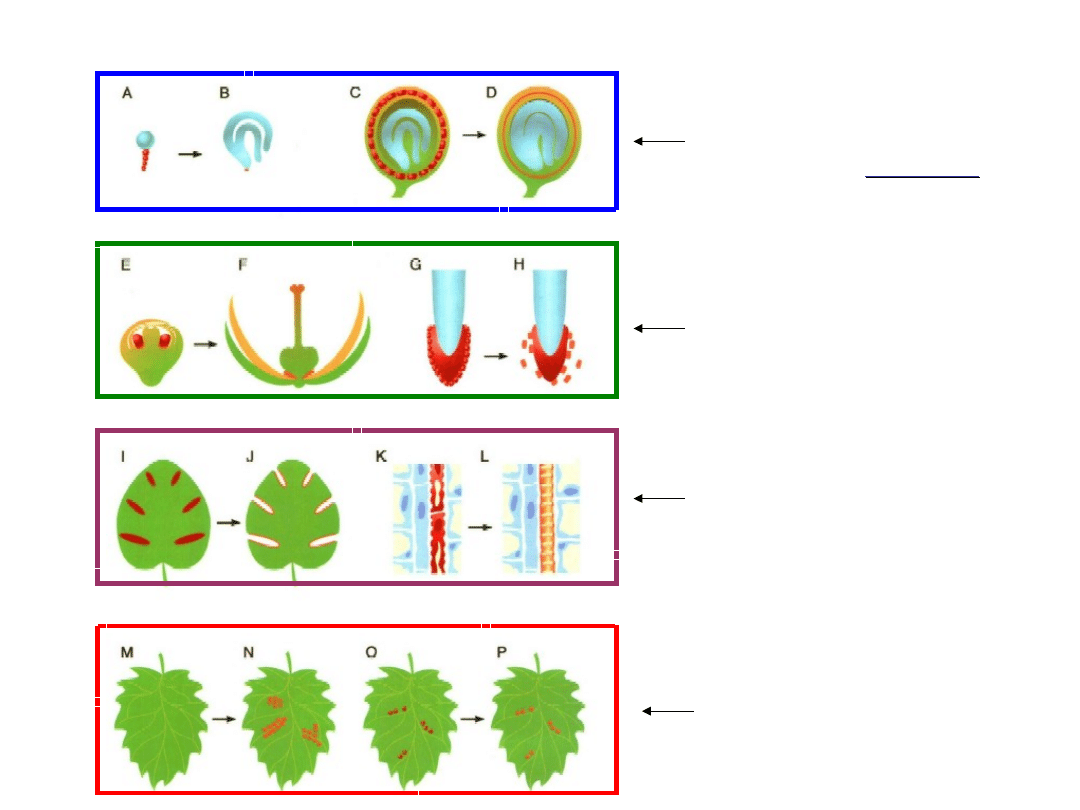
Funkcje PCD
Likwidacja komórek
zaatakowanych
przez patogeny
Likwidacja komórek
pełniących
czasowo
czasowo
określone funkcje
Likwidacja
niepotrzebnych komórek
Likwidacja komórek
podczas różnicowania
(specjalizacji) komórek

• W komórkach zwierzęcych PCD kończy się apoptozą.
Apoptoza to zaprogramowany genetyczne proces
obumierania komórek, w którego końcowej fazie
fragmenty struktury zostają
pochłonięte przez sąsiadujące
komórki (makrofagi) i całkowicie strawione w lizosomach.
• W komórkach roślinnych proces wchłaniania jest
niemożliwy
z powodu istnienia
ściany komórkowej.
U roślin podczas PCD następuje proces
samotrawienia AUTOFAGIA.
Różnica PCD w komórkach zwierzęcych
i roślinnych

PCD u roślin
AUTOFAGIA – ( gr. autos –sam, phagos – pożeracz) – polega
na trawieniu przez komórkę niektórych własnych
elementów jej struktury.
Może dotyczyć jednej komórki, bez włączania
sąsiadujących komórek.
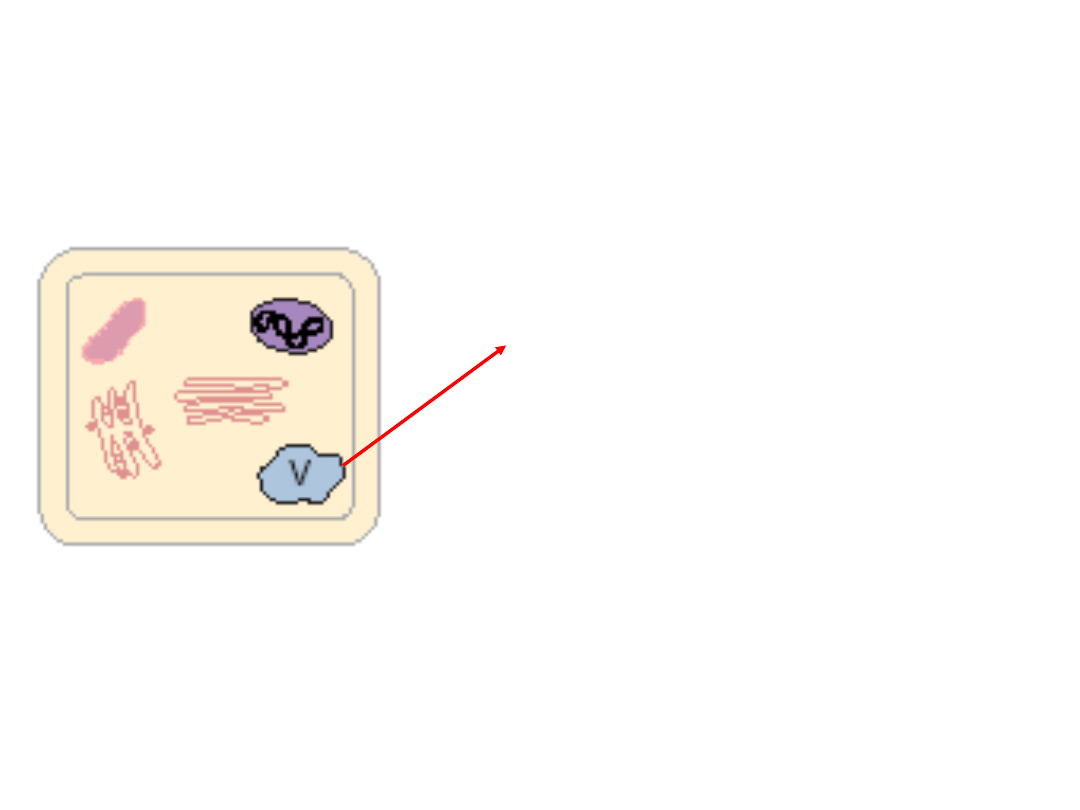
Autofagia
W procesie autofagii kluczową rolę
odgrywa
wakuola lityczna
,
prowadzącej do degradacji
protoplastu.
Zawiera ona zestaw enzymów
hydrolitycznych trawiących
komponenty komórkowe.
W komórce roślinnej może występować kilka
rodzajów wakuoli. Zróżnicowanie funkcji
wakuoli jest uwarunkowane m.in.
charakterem białek zgromadzonych w ich
wnętrzu.
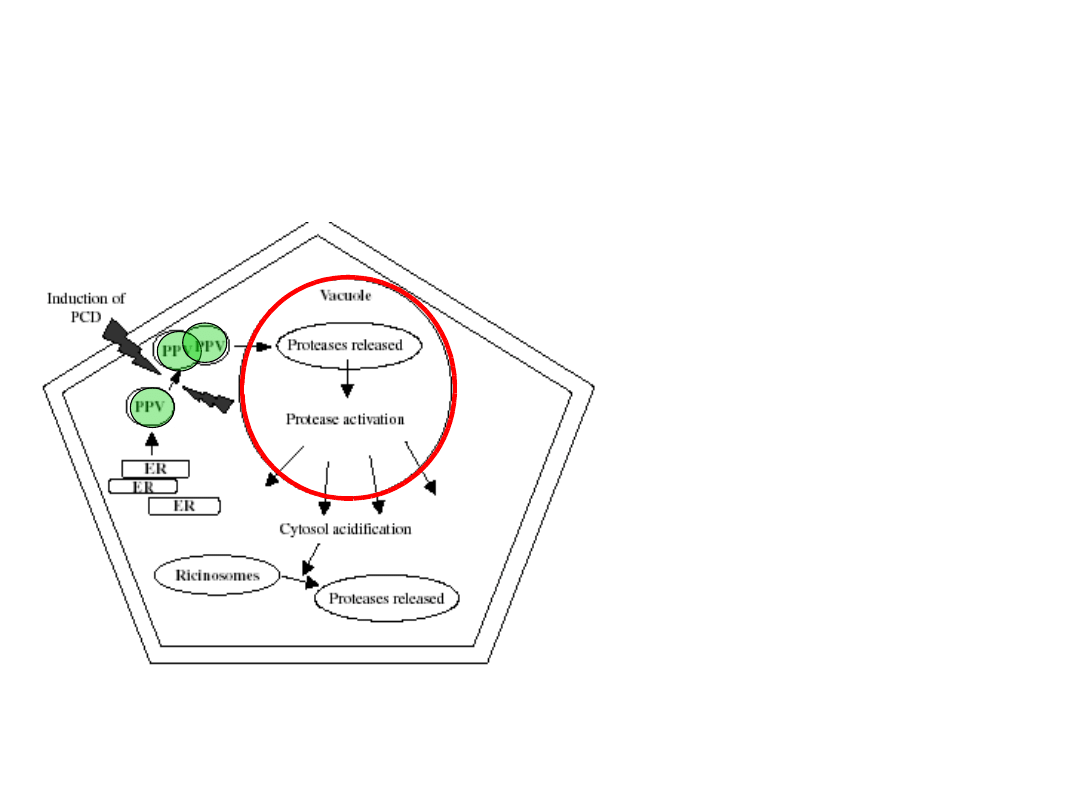
Wakuole lityczne
powstają, gdy w
soku wakuolarnym gromadzą
się liczne enzymy
hydrolityczne, co prowadzi do
znacznego zwiększenia jej
objętości.
Enzymy hydrolityczne,
kodowane przez genom
jądrowy lub pochodzące z ER,
syntetyzowane są w cytozolu i
dostarczane do wakuoli w
świetle
pęcherzyków
błonowych
(PPV)
jako
nieaktywne prekursory
proteaz. Wewnątrz wakuoli w
kwaśnym pH ulegają aktywacji.
Powstawanie wakuoli litycznych

Typy autofagii
Wakuola „wyposażona” w odpowiednie enzymy
podlega następnie kolejnym zmianom związanym z
procesem autofagii.
Trzy typy (ścieżki) autofagii:-
mikro
autofagia -
makro
autofagia
-
mega
autofagia
Mikro
autofagia
------
Mały fragment cytoplazmy
zostaje
„włączony” przez inwaginację (wpuklenie)
do wakuoli litycznej, powstaje ciało
autofagowe, które zostaje strawiony przez
proteazy.
Makro
autofagia
-----
Duży fragment cytoplazmy
wraz z
organellami zostaje „zamknięty” w
autofagosomie
(pęcherzyk otoczony
podwójną błoną).
Autofagosomy
powstaje prawdopodobnie z
ER.
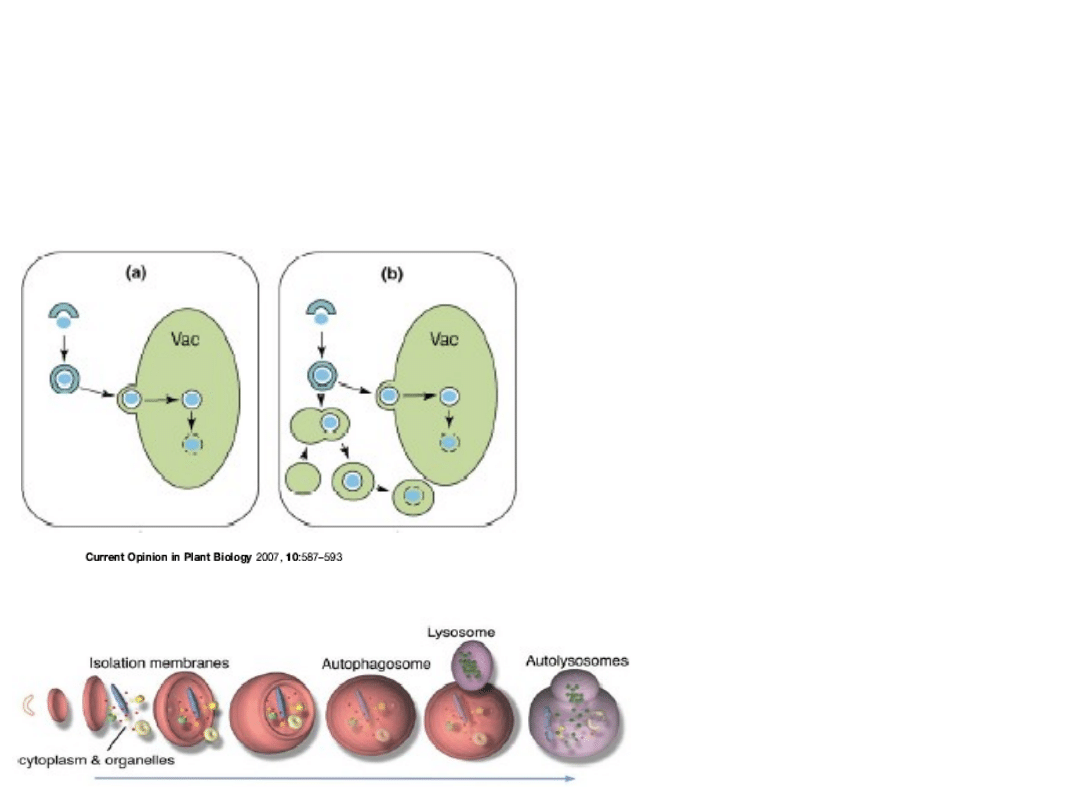
Makro
autofagia
(a) Na terenie cytoplazmy powstają
autofagosomy
które zostają
włączone do
wakuoli litycznych
.
Zewnętrzna błona
autofagosomu „zlewa” się z
tonoplastem, a autofagosom
przekształca się w ciało
autofagowe, stopniowo
degradowane przy udziale
wakuolarnych proteaz.
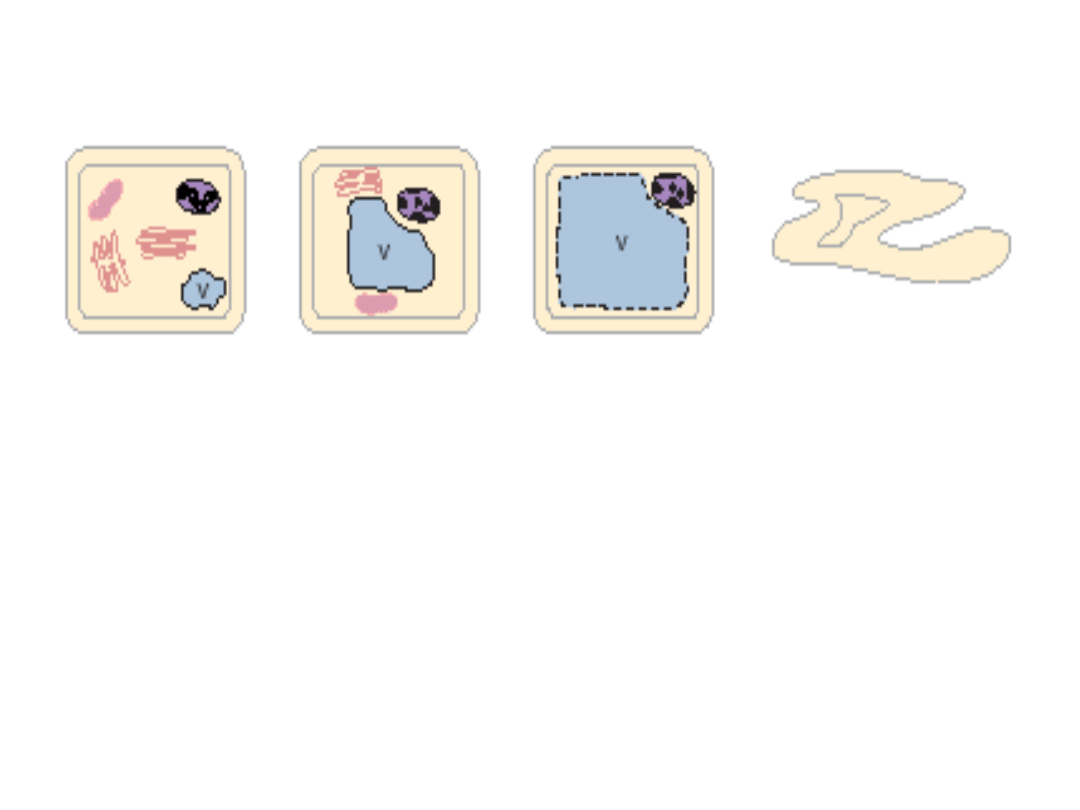
(b) U niektórych roślin (tytoń)
autofagosomy mogą także
zlewać się z lisosymami lub
endosomami na terenie
cytoplazmy. Powstają
autolisosomy
, na terenie
których autofagosomy mogą być
częściowo degradowane, zanim
zostaną włączone do wakuoli.
autofagosom
autolisosom
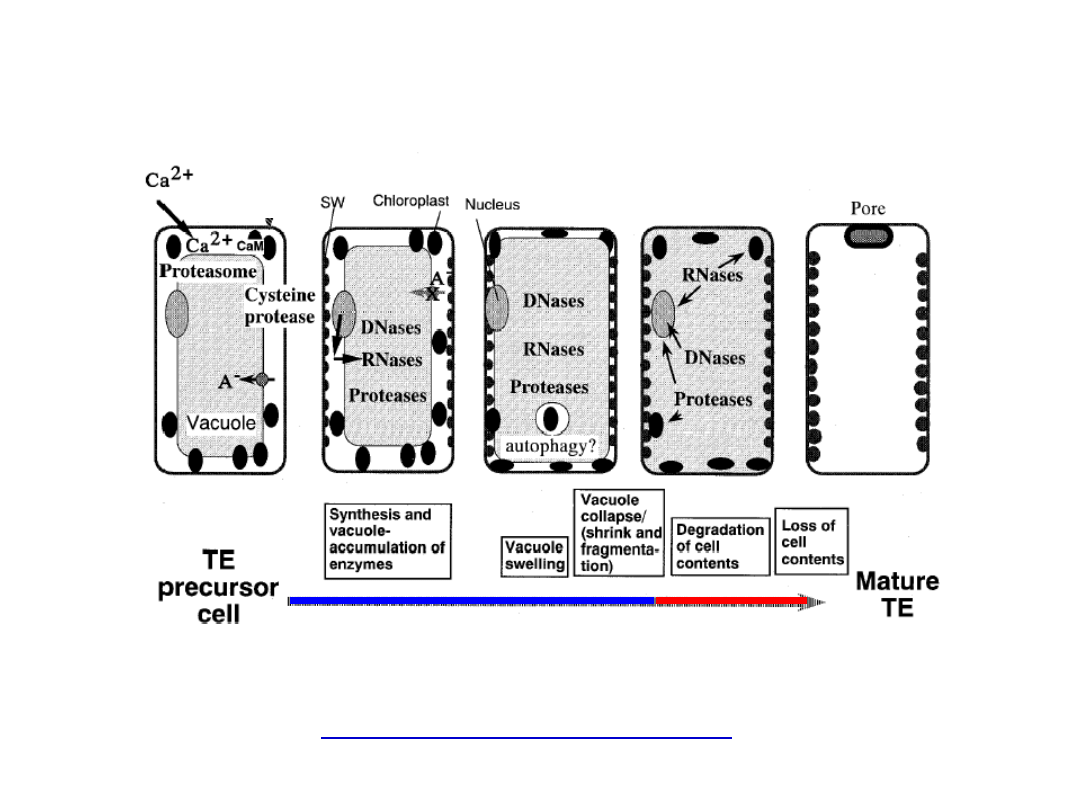
Mega
autofagia
1. Mega
autofagia rozpoczyna się od indukcji genów kodujących
specyficzne enzymy hydrolityczne, których ilość wzrasta w soku
wakuolarnym:
■
proteazy cysteinowe i serynowe
■ endonukleazy
(RNA-zy, DNA-zy)
■ kwaśne fosfatazy
■ lipazy
2. Wakuola znacznie się powiększa, zajmując niemal cały protoplast.
3. Następuje przerwanie tonoplastu i zakwaszenie protoplastu
4. Uwolnione enzymy rozpoczynają degradację składników komórki
PCD w roślinach
Obecnie wyróżnia się 2 typy przebiegu PCD w roślinach:
■
autofagia
- PCD z wytworzeniem autofagosomów
(mikro- i
makroautofagia)
■ apoptoza -
PCD bez ich wytwarzania autofagosomów
(=
megaautofagia)
■ nekrozy
- PCD nielizozomalna
Przebiegu PCD zależy od typu komórek/tkanek oraz czynników
środowiskowych i endogennych.

Rodzaje PCD w roślinach:
▲
PCD związana z rozwojem (autofagia)
-
obumieranie protoplastu podczas procesu
embriogenezy lub histogenezy
(powstawanie ksylemu, floemu,
aerenchymy) nazwano DCD (ang.
developmental cell death).
▲
PCD związana z reakcją na stresy (apoptoza)
(abiotyczne, biotyczne – HR, SAR)
PCD w roślinach
Warunkiem rozpoczęcia ksylogenezy jest odpowiednie
uformowanie wtórnej ściany komórkowej (zlignifikowanej).
Autofagia związana z rozwojem
(prowadząca do powstania naczyń ksylemu)
6 h
20 min.
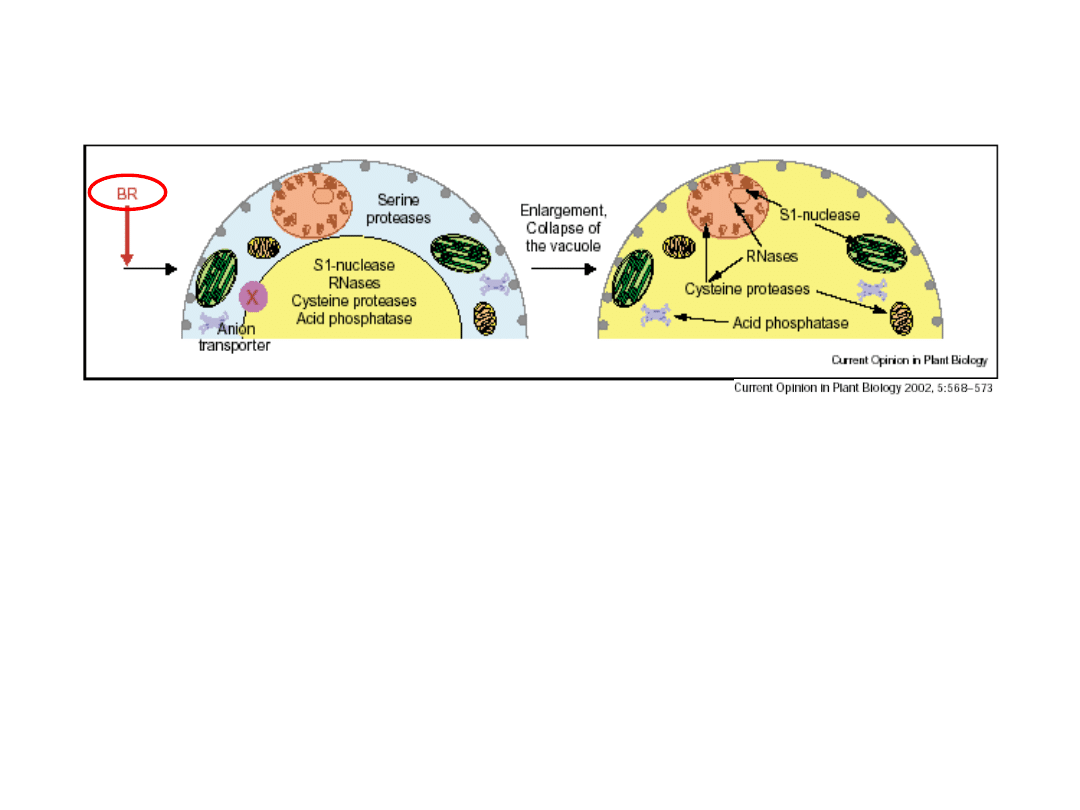
Brassinisteroidy
(+jony Ca)
indukują PCD związaną z
powstawaniem naczyń ksylemu:
■
indukcja formowanie
ściany wtórnej
komórek
■
stymulują syntezę i
akumulację w wakuoli
specyficznych enzymów
hydrolitycznych:
S1-
nukleazy, DNA-zy, RNA-zy,
proteazy cysteinowe,
kwaśne fosfatazy.
■
pęknięcie wakuoli
powoduje wypłynięcie soku
wakuolarnego wraz z
enzymami
■
następuje
szybka
degradacja organelli
komórkowych
■
częściowej
degradacji ulega także
ściana komórkowa (powstają
pory
naczyń ksylemu)
Autofagia związana z rozwojem
(prowadząca do powstania naczyń ksylemu)
Ca

Kluczowym momentem PCD jest przerwanie tonoplastu,
dalsze etapy trwają ok. 10-20 min
Sekwencja zdarzeń od momentu przerwania ciągłości
tonoplastu:
• degradacja zawartości chloroplastów (DNA),
• kondensacja chromatyny w jądrze komórkowym,
• stopniowa degradacja chromatyny, otoczka jądrowa
pozostaje nienaruszona do późnych etapów PCD.
Autofagia związana z rozwojem
(prowadząca do powstania naczyń ksylemu)
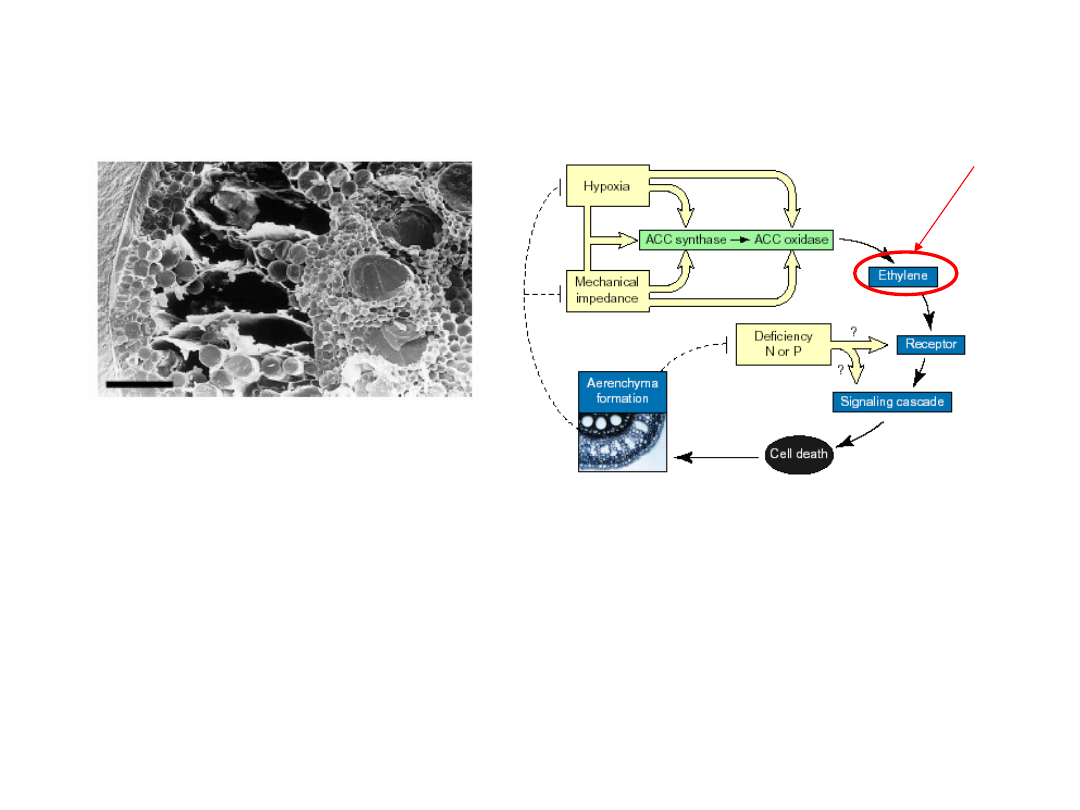
Adaptacji korzeni do niekorzystnych warunków środowiska
(hypoksja) jest regulowana przez etylen i ROS.
Wykształcenie aerenchymy umożliwia dyfuzję tlenu i
przebieg procesów metabolicznych funkcjonujących w
warunkach tlenowych
ROS
Powstawanie
aerenchymy jest
związane z
całkowitym
zanikiem ścian
wybranych komórek.
Autofagia związana z rozwojem
(prowadząca do powstania aerenchymy)
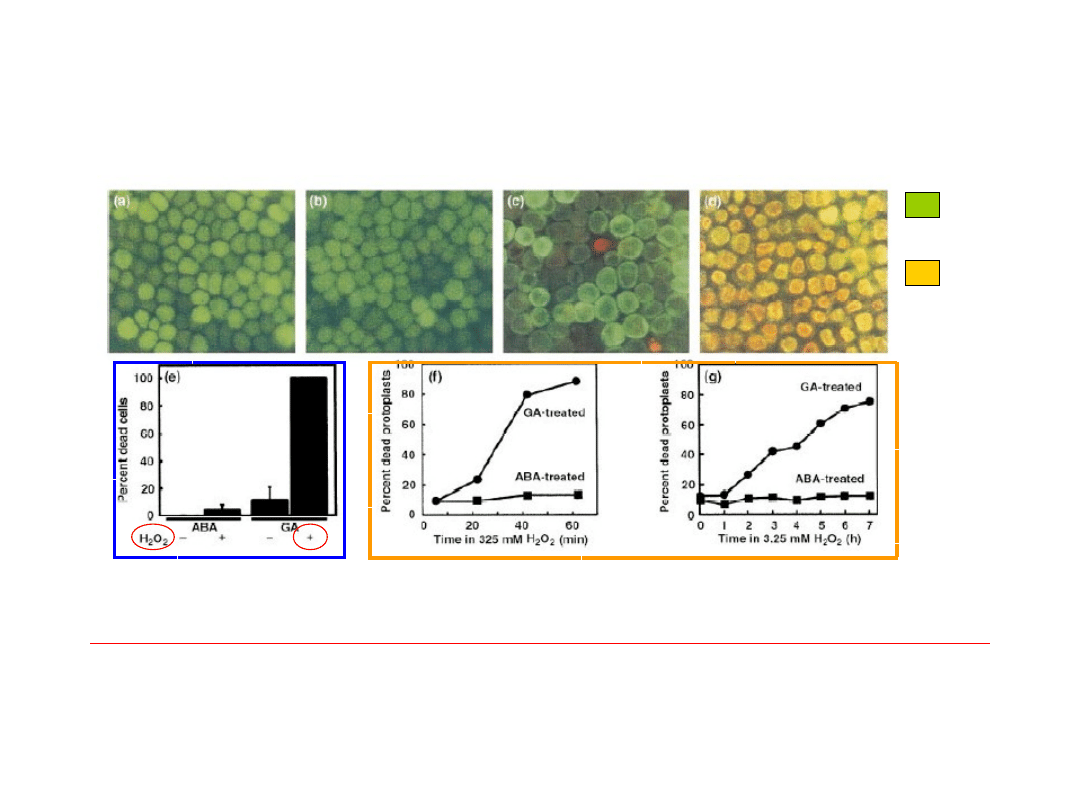
Wpływ
ROS
i fitohormonów na
PCD komórek
(a-e) i
protoplastów
(f-g) w warstwie aleuronowej kiełkującego jęczmienia.
GA – stymulują aktywność enzymów hydrolizujących materiały zapasowe,
ROS – odgrywają ważną rolę w degradacji komórek, powodując uszkodzenia składników komórkowych.
+ABA + H
2
O
2
+GA
- k. żywe
- k.
martwe
+ABA
+GA+H
2
O
2
Autofagia związana z rozwojem
(degradacja warstwy aleuronowej podczas kiełkowania
zbóż)
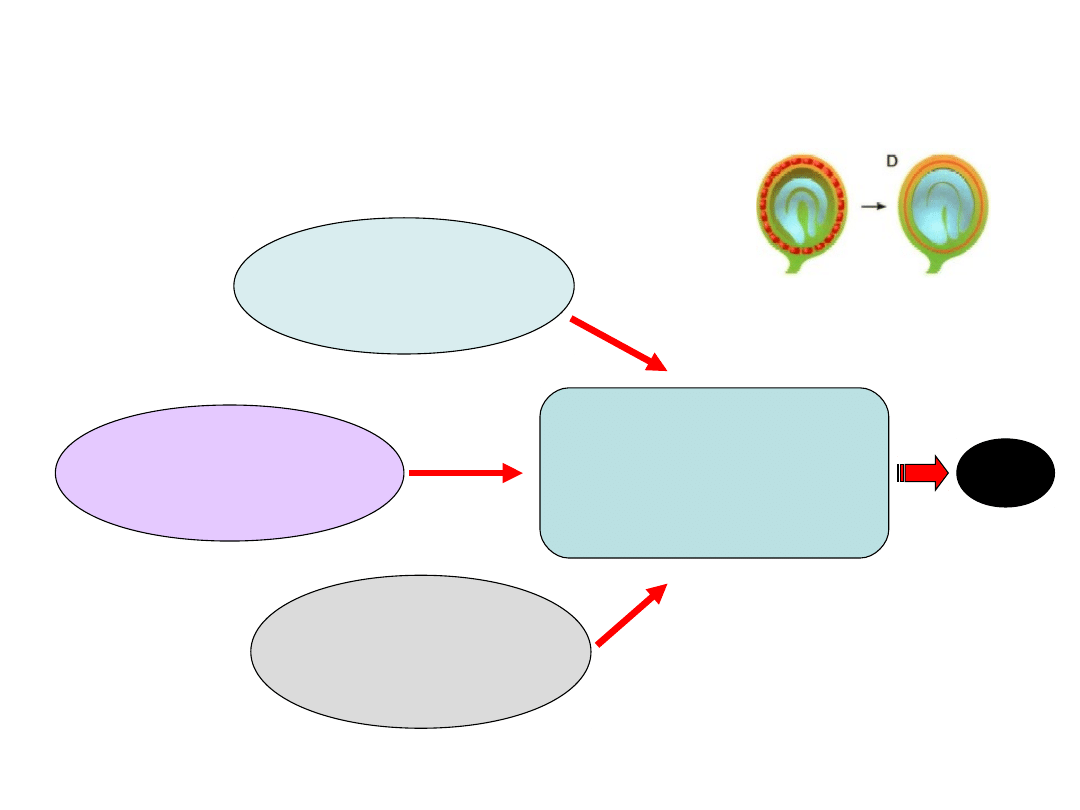
Produkcja i akumulacja
ROS
Spadek aktywności
mitochondriów
(deficyt ATP, słaba
regeneracja anty-
oksydacyjna
Obniżenie aktywności
systemu antyoksydacyjnego
(SOD, CAT, APOX)
Oksydacyjne uszkodzenia
błon
(peroksydacja lipidów,
degradacja białek
i rozpad plazmalemy)
PCD
Autofagia związana z rozwojem
(udział
ROS
w degradacji warstwy aleuronowej podczas
kiełkowania zbóż)
Apoptoza podczas reakcji nadwrażliwości
(HR) w odpowiedzi na atak patogenów
• Reakcja nadwrażliwości to bardzo
szybka reakcja
, bez
wytwarzania autofagosomów,
podobna do apoptozy (u
zwierząt),
• Pierwsze symptomy HR następują w
mitochondriach
-
wypływ cytochromu c.
• Wczesne zmiany w jądrze komórkowym, przed
pęknięciem wakuoli
• Szybkie obkurczanie cytoplazmy
• Produkcja dużej ilości ROS
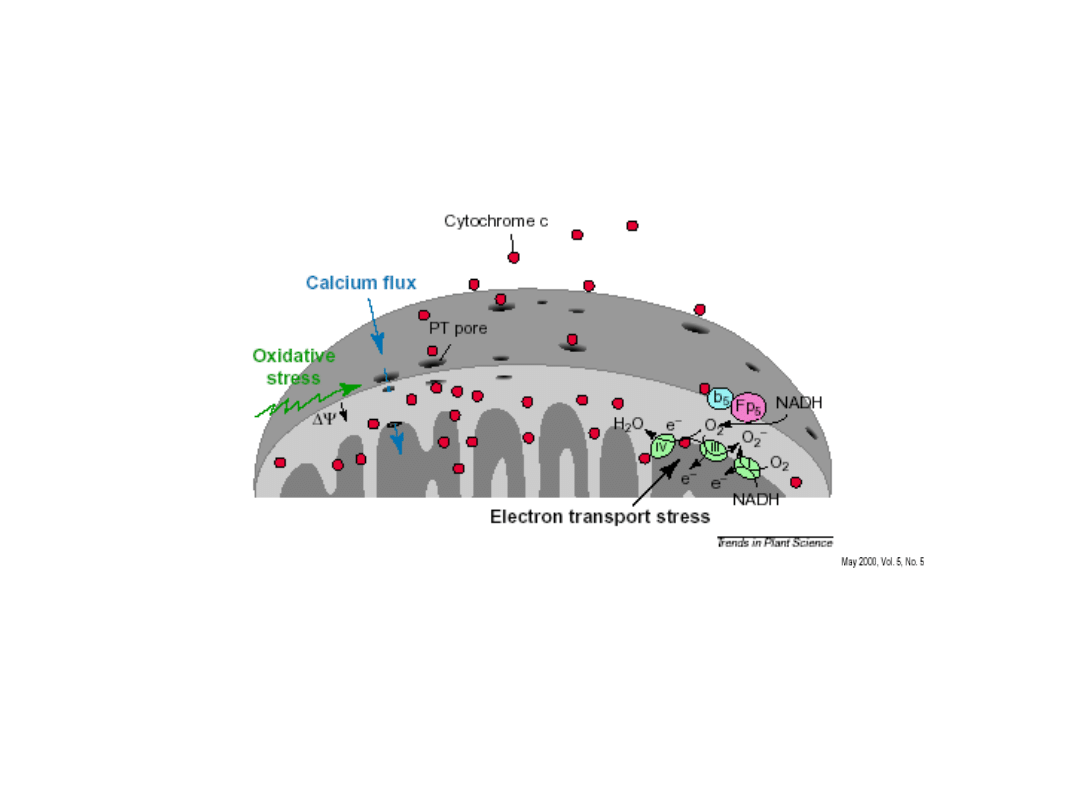
PCD spowodowana atakiem patogenu
Pierwsze symptomy HR następują w
mitochondriach
(wypływ cytochromu c)
ROS -
powstające podczas reakcji nadwrażliwości (HR) wraz z
białkami BAX powodują powstanie
MEGAKANAŁÓW
w
zewnętrznej błonie mitochondrialnej, pozwalające na przejście
białek (m.in. cytochromu c) do cytosolu.
Białka te stymulują
kaspazy
i inne pomocnicze białka
apoptotyczne, co aktywuje nieodwracalną PCD.
Mechanizm PCD
spowodowana atakiem
patogenu
Odpowiedź
systemiczna
Odpowiedź rośliny na patogeny może być:
• miejscowa
• systemiczna (w miejscu oddalonym od miejsca ataku
patogena)
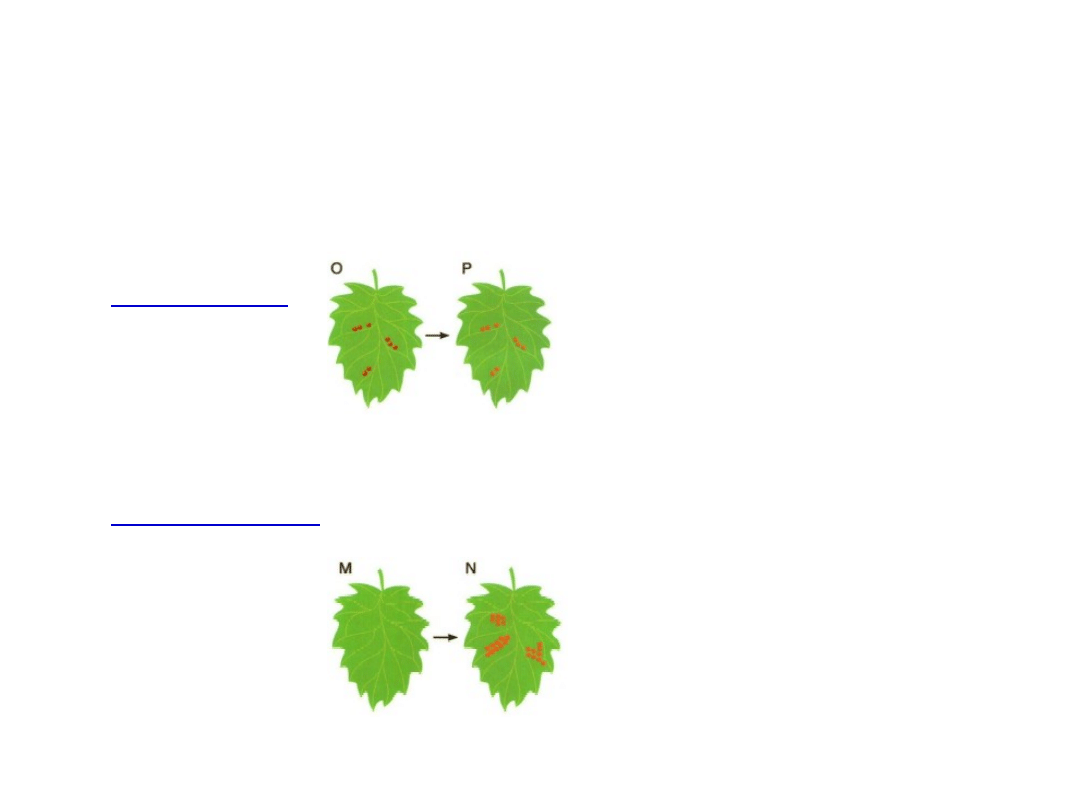
Odpowiedź miejscowa

Nekrozy
(odpowiedź miejscowa)
• Powstanie nekroz polega na
zahamowaniu
w protoplaście
obumierającej komórce podstawowych
procesów
biosyntetycznych
, któremu towarzyszy dezorganizacja
błon, bez udziału litycznej wakuoli i autofagosomów.
Z
reguły dotyczy małej liczby komórek
.
• Szybka reakcja na atak patogenu,
wskutek nagromadzeniu
ROS
.
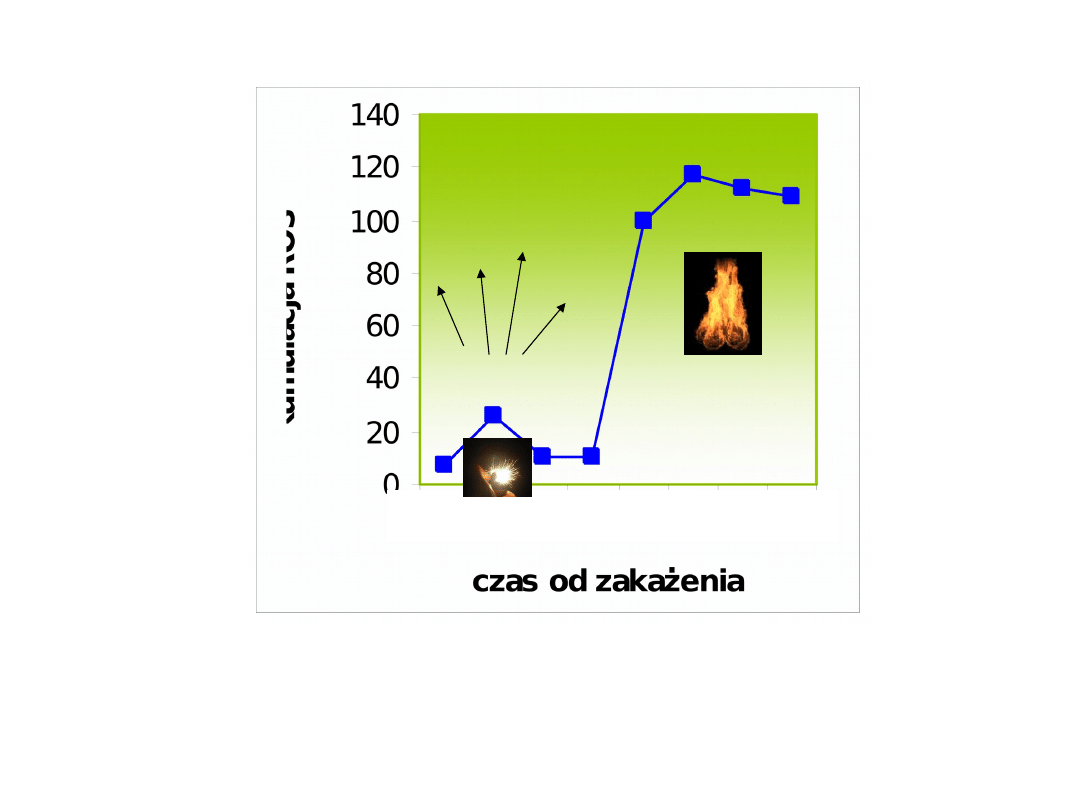
Dwufazowa akumulacja ROS podczas
HR
0 1 2 3 4 5 6 7
1.
sygnał
2.
fala
oksydacyj
na
ROS (H
2
O
2
) i RNS (NO) biorą udział we wczesnych etapach
odpowiedzi na patogen:
1. jako sygnał, „iskra” uruchamiają
ścieżki obrony,
2. jako fala oksydacyjna powodują nekrozy lub PCD,
które
ograniczają rozprzestrzenianie patogenu.
NEKROZY
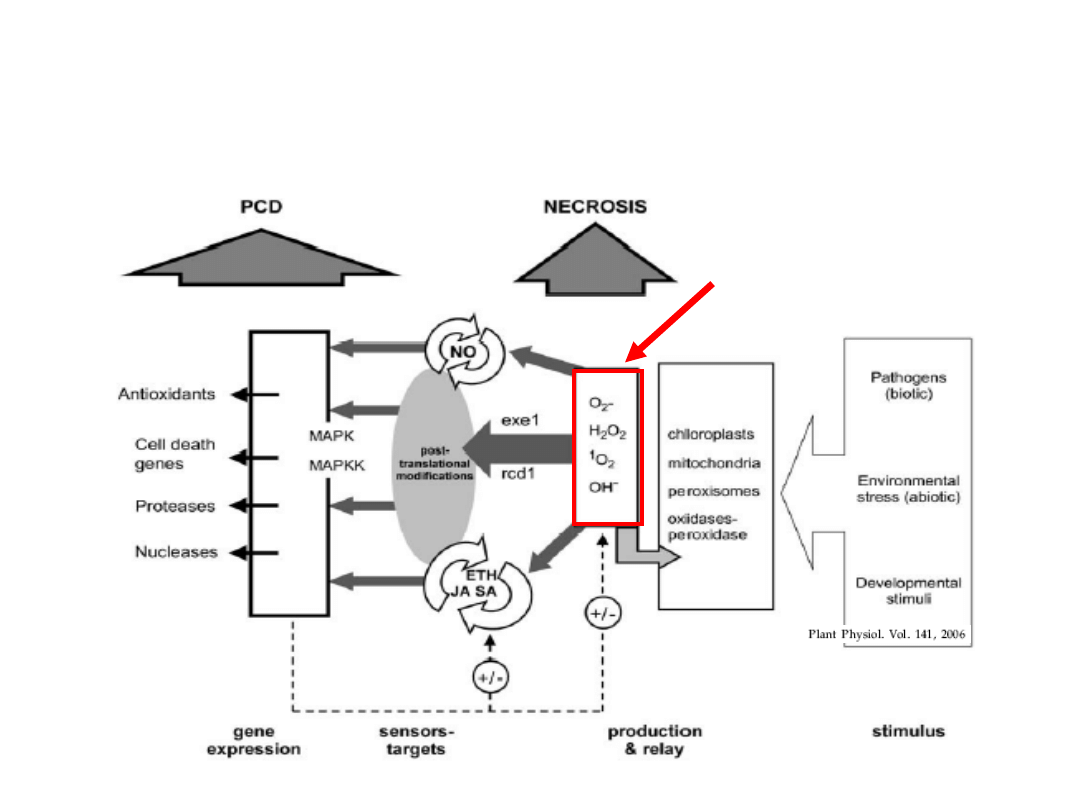
Warunki środowiska i faza rozwojowa
kształtują odpowiedź morfogenetyczną
rośliny
ROS = sygnał ”śmierć”

PCD podczas
starzenia
■ Proces obumierania komórek podczas starzenia jest znacznie
dłuższy
niż PCD
związana z rozwojem czy atakiem patogena.
■ Starzenie jest bowiem
poprzedzony redystrybucją materiałów
zapasowych
do
innych organów rośliny (młodsze liście, kwiaty i rozwijające się owoce i nasiona).

PCD podczas
starzenia
■
PCD w trakcie starzenia charakteryzują w pierwszej
kolejności
szybkie zmiany
w chloroplastach
w chloroplastach
- zahamowanie syntezy białek poprzez zahamowanie
aktywności polimerazy RNA,
- degradacja tylakoidów,
- degradacja białek i DNA w stromie).
■ Proces starzenia może być
odwrócony
.
Przywrócenie aktywności polimerazy RNA może prowadzić
do zatrzymania degradacji chloroplastów, powtórnego ich
funkcjonowania i zazielenienia liści.
■
Dopiero gdy proces obumierania
chloroplastów
stanie się
nieodwracalny rozpoczynają się zmiany w wakuoli.
■
Zmiany w jądrze zachodzą na końcu procesu PCD
(kondensacja chromatyny, fragmentacja DNA przy udziale
swoistych endonukleaz).

Regulacja starzenia
Proces starzenia jest regulowany przez:
■
czynniki egzogenne
(stres, brak
substancji pokarmowych, ciemność),
■
endogenne
(wiek, hormony)
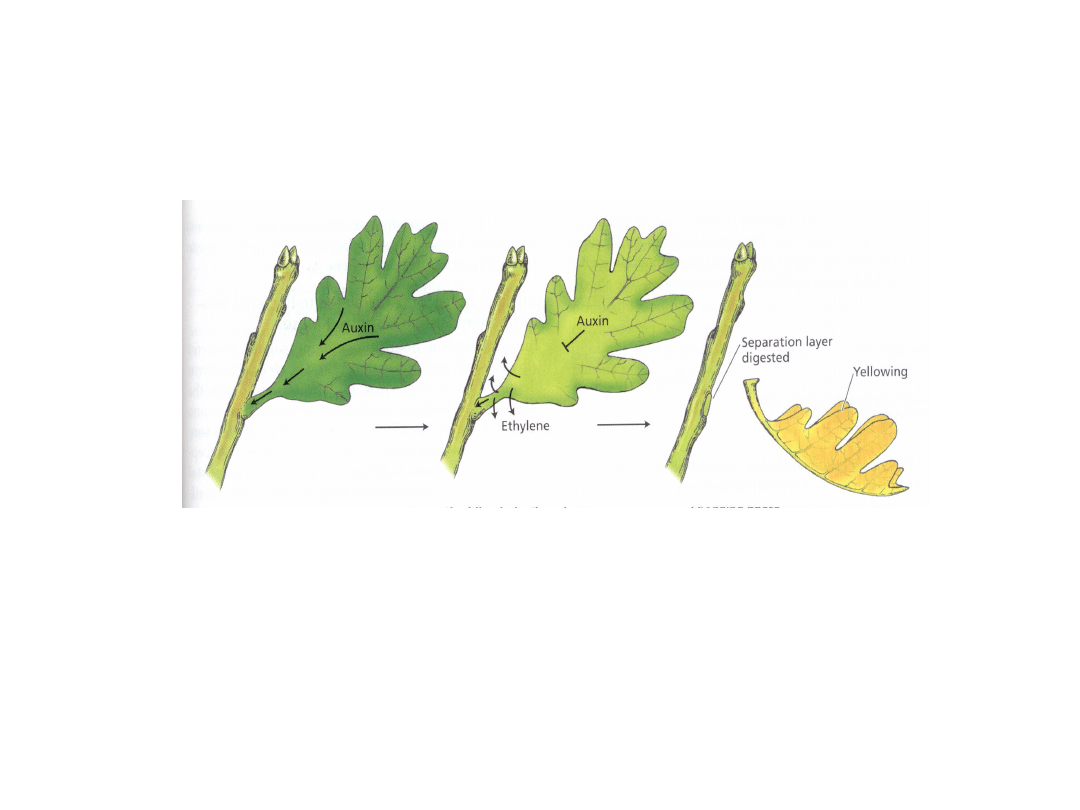
Etylen indukuje powstawanie warstwy
odcinającej
Wysokie stężenie
auksyn hamuje
syntezę etylenu
Spadek stężenia
auksyny
stymuluje
syntezę etylenu
Etylen (wraz z ABA)
stymuluje degradację
ścian komórkowych przy
udziale enzymów
hydrolitycznych (celulazy,
poligalakturonazy)
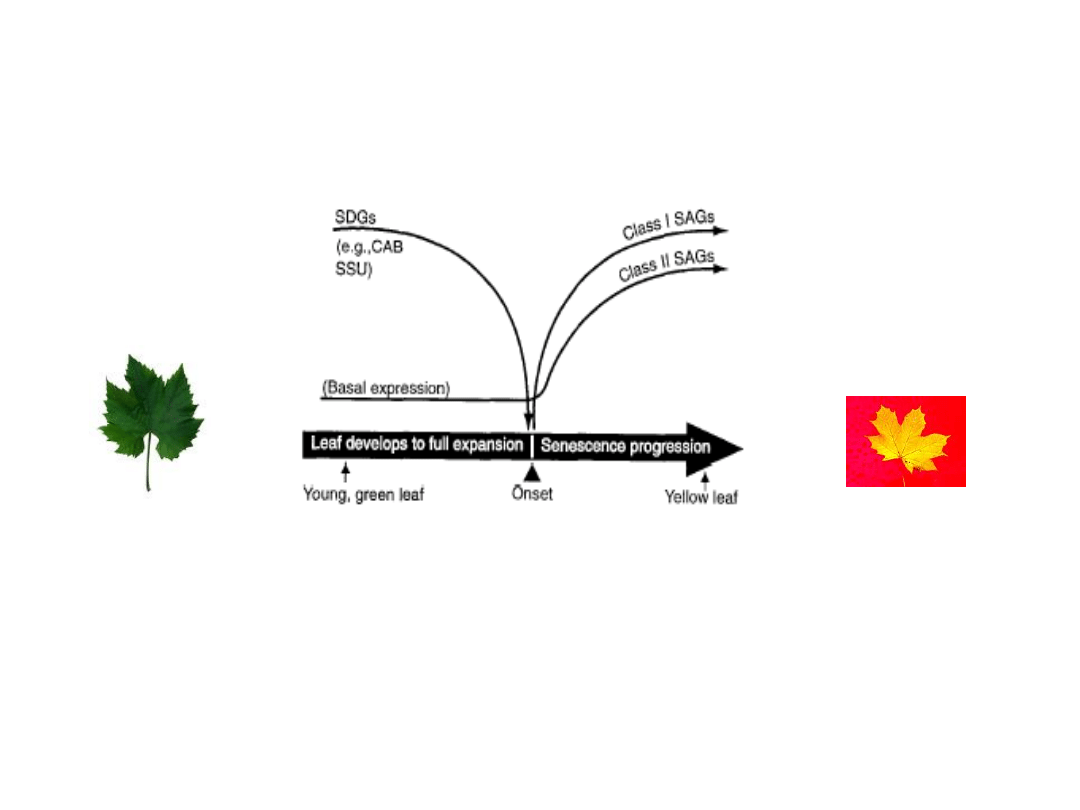
Starzenie liści zależy od ich wieku
i jest regulowane przez specyficzne geny: SDG
i SAG
Ekspresja genów podczas rozwoju liści (SDG – geny
hamowane podczas starzenie) i ich starzenia (SAG – geny
stymulowane podczas starzenie)
SDGs – senescence-decreased genes
SAGs – senescence-associated genes
Ekspresja genów SAG jest stymulowana przez szereg hormonów (JA, ABA, etylen)
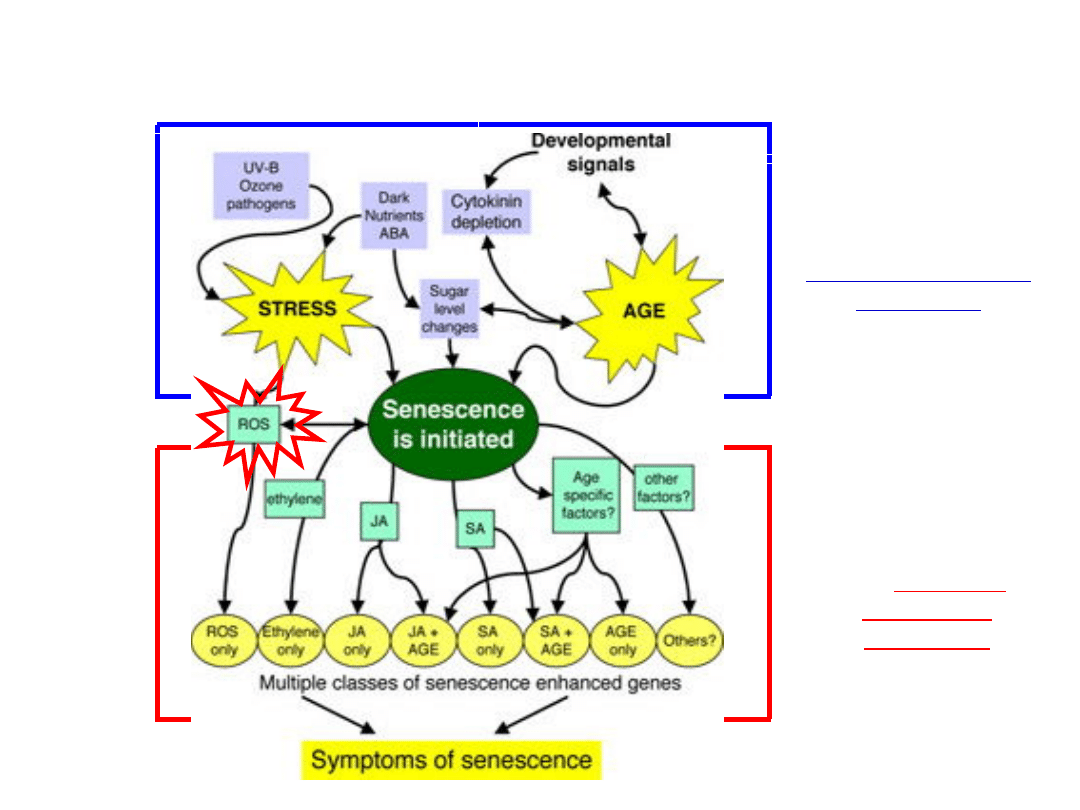
Regulacja procesu starzenie roślin
Endo- i
egzogenne
sygnały inicjacją
starzenia
ROS
oraz SA i
hormony (etylen,
JA) regulują
realizację
starzenia
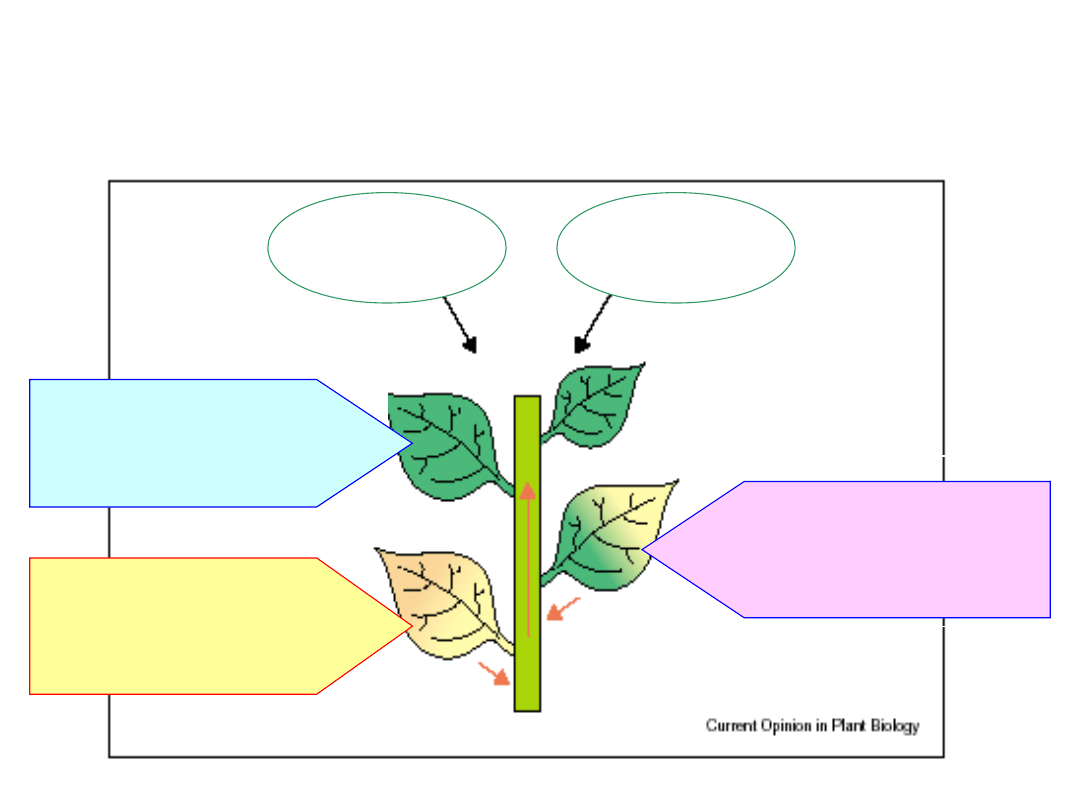
I. Faza inicjacji:
- spadek intensywności fotosyntezy
- zmiana wzorca dystrybucji asymilatów
- zatrzymanie proc. metabolicznych
II. Faza degradacji
- degradacja składników komórkowych
białek, lipidów, kw. nukleinowych
- transport produktów rozpadu do
innych organów
III. Faza końcowa
-
wzrost produkcji ROS
- degradacja jądra i mitochondriów
- utrata integralności komórki
- śmierć komórki
Sygnał endogenny
rozwojowy
Sygnał egzogenny
środowiskowy
Przebieg procesu starzenia
(fazy starzenia)
Podsumowanie
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
Wyszukiwarka
Podobne podstrony:
Programowana śmierć komórki w aspekcie posprzętnej trwałości kwiatów
Programowa śmierć komórki apoptoza
15.Programowana śmierć komórki, studia-biologia, Opracowane pytania do licencjatu
SKRÓCONY PROGRAM OCHRONY ROŚLIN SADOWNICZYCH PRZED CHOROBAMI
Poliaminy w programowanej śmierci komórki
WZROST, RÓŻNICOWANIE, STARZENIE SIĘ I NATURALNA ŚMIERĆ KOMÓREK
SKRÓCONY OPIS PROGRAMU PSPICE
2014 11 26 „Ignis Ardens W 100 lecie śmierci Św Piusa X” Program
ŚMIERĆ I JEJ OZNAKI
w4 skrócony
w3 skrócony
Nowy Prezentacja programu Microsoft PowerPoint 5
STARZENIE LEK
więcej podobnych podstron