
Witam w kolejnym opracowaniu wykładów z Fizjologii i anatomii, tym razem omówimy różne
rodzaje mięśni. Zalecam jednak zapoznać się z właściwościami tkanki mięśniowej, bo prof.
przeszedł od razu do rzeczy.
Podział mięśni:
-Mięśnie szkieletowe (poprzecznie prążkowane)
-Mięśnie gładkie
-Mięsień sercowy
Mięśnie szkieletowe:
-Otrzymują pobudzenie od układu nerwowego, w miejscu zwanym synapsą nerwowo-mięśniową.
-Synapsa nerwowo-mięśniowa jest synapsą chemiczną, której częścią presynaptyczną jest
komórka nerwowa, a częścią postsynaptyczną komórka mięśniowa. Rolę przekaźnika pełni
acetylocholina. Działanie tej synapsy wygląda dokładnie tak samo jak działanie synapsy chemicznej
omówionej w poprzednim opracowaniu.
-Acetylocholina: jest przekaźnikiem pobudzającym – wywołuję depolaryzacje błony
postsynaptycznej, poprzez łączenie się z receptorem nikotynowym (mechanizm ligandozależny). To
połączenie otwiera kanał sodowo-potasowy. Potas zaczyna wypływa z komórki, a sód napływać.
Napływ jonów sodowych jest jednak większy niż odpływ jonów potasowych, dzięki czemu
dochodzi do depolaryzacji błony. Potencjał wywołany przez acetylocholinę zawsze wywoła
potencjał czynnościowy (zawsze przekroczy wartość progową) Jest to niezwykle ważne z punktu
widzenia odruchów obronnych np. obrony przed bólem. Poza tym dzięki tej pewności wzbudzenia
potencjału czynnościowego mamy również pewność, że zawsze wykonamy ruch.
A teraz mały spinoff
Warunki jaki musi spełniać jakaś substancja, żeby być przekaźnikiem.
–
posiadanie agonisty i antagonisty
–
agonista – to taka substancja, która działa podobnie do przekaźnika. Ma inną budowę, ale
wywołuje ten sam skutek, czyli otwarcie kanału jonowego. Agonistą dla acetylocholiny jest
nikotyna
–
Antagonista – to substancja mająca przeciwne działanie do przekaźnika – powoduje
zamknięcie kanału jonowego. Antagonistą dla acetylocholiny jest kurara – substancja
stosowana przy operacjach na mięśniach (powoduje ich wiotczenie)
–
Posiadanie receptora – receptorem dla acetylocholiny jest receptor muskarynowy (a także
nikotynowy)Kiedy acetylocholina łączy się z receptorem muskarynowym, agonistą w
stosunku do niej jest atropina (która nie łączy się z receptorem nikotynowym)
Koniec spinnoffu
Potencjał czynnościowy na komórce mięśniowej jest konieczny do wywołania skurczu. Aby
zrozumieć mechanizm, trzeba poznać podstawową jednostkę czynnościową mięśnia, czyli
sarkomer.
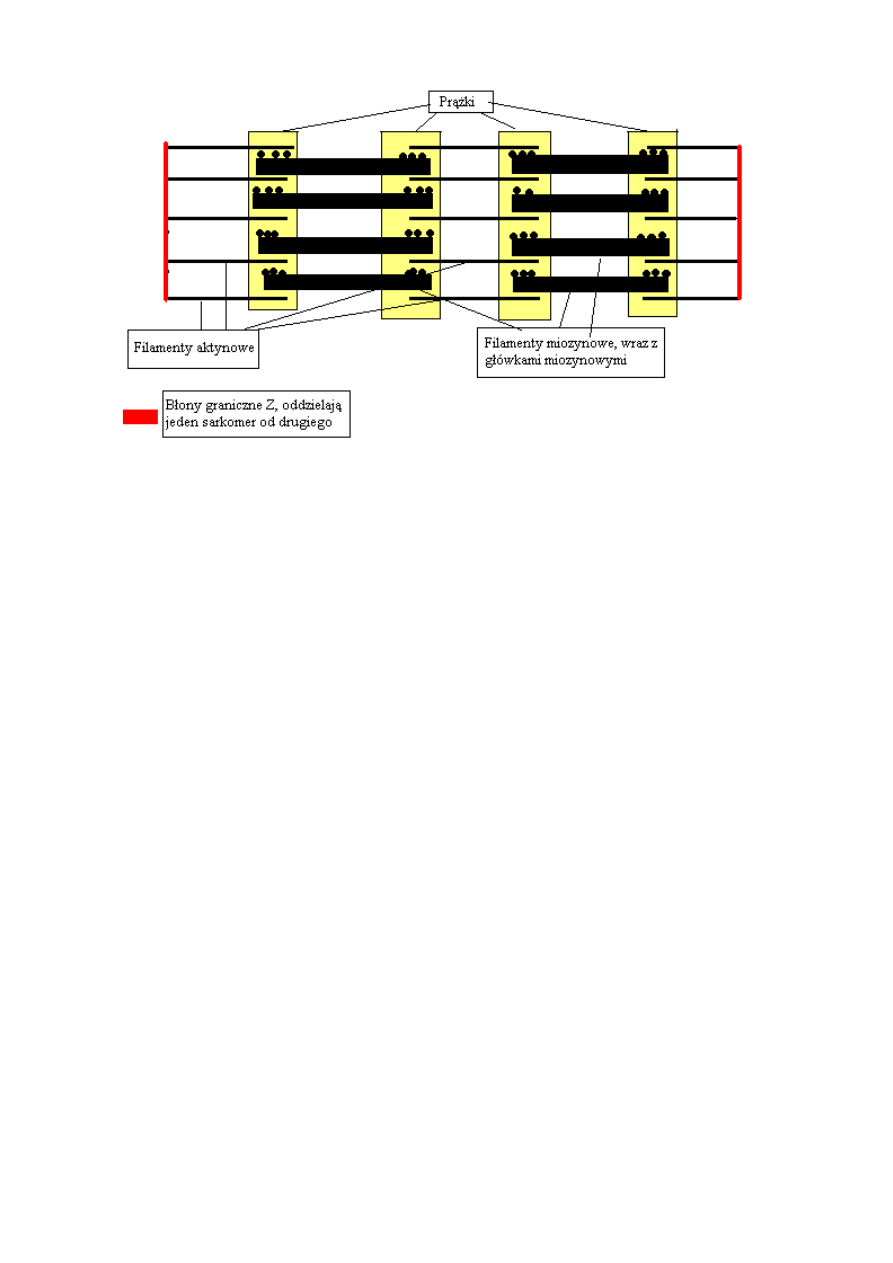
Sarkomery ułożone są szeregowo (jeden za drugim) i budują miofibryle. Sarkomer składa się z
dwóch błon granicznych Z, które oddzielają go od innych sarkomerów. Do błon przyczepione są
filamenty aktynowe, pomiędzy którymi znajdują się filamenty miozynowe. (stąd biorą się prążki)
Molekularny mechanizm skurczu
Aktyna i miozyna nie są ze sobą połączone, należy więc wytworzyć fizyczne połączenie między
nimi. Miozyna posiada główki miozynowe, natomiast aktyna posiada odpowiednie miejsce do ich
przyczepu. Przy spoczynku mięśnia miejsce na aktynie jest zasłonięte przez kompleks tropinowo-
tropomiozynowy. Odsłonięcie tego miejsca jest warunkowane obecnością jonów Ca
2+
. Jony te łączą
się z tropiną, co powoduje konformacyjną zmianę przestrzenną kompleksu, a w rezultacie
odsłonięcia miejsca przyczepu dla miozyny.
Przyłączenie miozyny do aktyny jest dopiero początkiem skurczu. Teraz należy wsunąć miozynę
pomiędzy aktynę. Główka miozynowa ma charakter ATPazy, czyli może uwalniać energię z ATP.
Tutaj po raz kolejny dają o sobie dać jony wapniowe, które aktywują ATPazę. Energia uwolniona z
hydrolizy ATP sprawia, ze główka miozynowa się skręca, a to owocuje wsunięciem się miozyny
między filamenty aktynowe.
Podsumowując: Aby skurcz był możliwy potrzebne nam są jony wapniowe (Ca
2+
.), które
odblokują miejsce przyczepu miozyny na aktynie i wyzwolą energię potrzebną do skręcenia główek
miozynowych.
Skąd zatem wziąć te jony?
–
w cytoplazmie jest ich mniej niż na zewnątrz komórki. Potencjał czynnościowy otwiera
kanały jonowe, poprzez które wnikają one do wnętrza komórki
–
w komórkach mięśniowych znajdują się sarkoplazmatyczne magazyny jonów wapniowych,
występujące w formie cystern, zaopatrzonych w kanały jonowe bramkowane napięciem, z
których w razie potrzeby również te jony są uwalniane.
Rodzaje skurczów:
–
pojedyncze, które dzielą się na: izotoniczne i izometryczne o auksotoniczne
–
złożone, które dzielą się na zupełne i niezupełne
Skurcz pojedynczy to taki, który wywoływany jest przez pojedynczy bodziec
Skurcz izotoniczny, to taki przy którym zmienia się długość mięśnia, lecz nie zmienia się jego
napięcie
Skurcz izometryczny to taki, przy którym zmienia się napięcie mięśnia, lecz nie zmienia się jego
długość
Skurcz auksotoniczny to taki przy którym zmienia się i długość i napięcie mięśnia
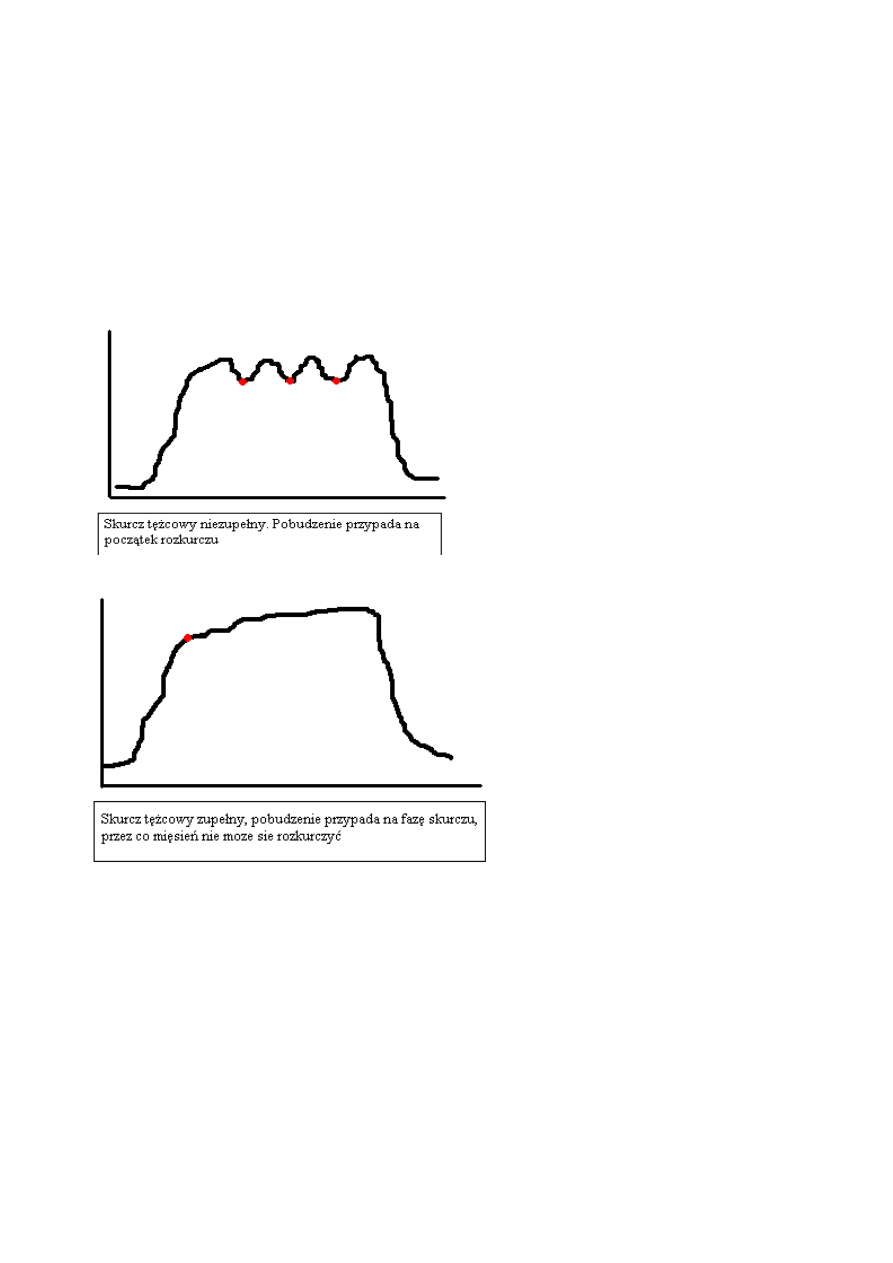
Skurcz złożony (tężcowy) to taki, który wywołany jest przez kilka nakładających się bodźców
Skurcz złożony zupełny – występuje w
ówczas, gdy pobudzenie następuje zanim mięsień
zacznie się rozkurczać
Skurcz złożony zupełny
powstaje przy pobudzenia mięśnia w okresie rozkurczu
Wykład sporządził Maciej „Jelcyn” Gawroński. Sponsorzy: Mictosoft Paint, Open office, tabaka
Gletscherprise i literka B jak „Bad Motherfucker!”
Wyszukiwarka
Podobne podstrony:
Fizjologia i Anatomia wyklad I
Fizjologia i Anatomia wyklad VIII
Fizjologia i Anatomia wyklad VII
Anatomopatologia Wykłady III Semestr
Fizjologia i anatomia wyklad IV
Fizjologia i Anatomia wyklad I
Fizjologia i anatomia człowieka Krew (wykłady i ćwiczenia)
fizjologia roślin - wykład (8.01.2014), Semestr III, Fizjologia Roślin, Wykłady
Fizjologia - wykład III - 12.03.2013, Ogrodnictwo 2011, Fizjologia roslin
fizjologia roslin wyklad, Rok III, Rok II, Semestr III, Fizjologia roslin
Fizjologia wyklad III, SGGW, fizjologia zwierzat
Fizjologia roślin (18.12.2013), Semestr III, Fizjologia Roślin, Wykłady
Fizjologia roślin - wykład (27.11.2013), Semestr III, Fizjologia Roślin, Wykłady
Fizjologia roślin - wykład (04.12.2013), Semestr III, Fizjologia Roślin, Wykłady
TBL WYKŁAD III Freud
wykład III Ubezpieczenia na życie2011
więcej podobnych podstron