
BŁONY BIOLOGICZNE

Błony biologiczne występują we wszystkich
znanych układach biologicznych zdolnych do
samodzielnego życia. Oddzielają one
komórkę od środowiska, a w komórkach
Eukariota dzielą również wnętrze komórki na
mniejsze obszary o zróżnicowanych
funkcjach (budują struktury błoniaste:
endoplazmatyczne retikulum, aparat
Golgiego, pojedyncza błona otacza wakuolę,
lizosomy, peroksysomy a podwójna jądro
komórkowe, mitochondria i plastydy).
Błony różnią się składem białek i fosfolipidów
oraz nieznacznie właściwościami.

Błony biologiczne uczestniczą w:
• biernym lub czynnym, selektywnym
transporcie jonów i substancji niejonowych,
• wydzielaniu produktów komórki do
środowiska (egzocytoza) oraz pobieraniu
makrocząsteczek do komórki (endocytoza),
• reakcjach na sygnały pochodzące ze
środowiska (transdukcja sygnałów) poprzez
receptory błonowe,
• przenoszeniu sygnałów do innych okolic
komórki lub przekazywaniu ich do innych
komórek,
• oddziaływaniu między komórką i podłożem
oraz między komórkami.

Ich rolą jest też:
• oddzielenie wnętrza komórki od środowiska,
• oddzielanie w komórkach kompartymentów
(przedziałów) o różnej koncentracji różnych
substancji (enzymów, jonów, substratów),
• pośredniczenie w transporcie biernym i
czynnym,
• wytwarzanie potencjału elektrochemicznego
- różnej koncentracji jonów,
• miejsce przebiegu procesów (np. łańcuch
transportu elektronów w mitochondriach i
chloroplastach)

Teorie budowy błon
1. Model lipidowy - W roku 1895 Overton opierając się na fakcie,
że substancje rozpuszczalne w tłuszczach wnikały do komórki
bardziej efektywnie niż nierozpuszczalne - wydedukował, że lipidy
muszą stanowić ważny składnik błony plazmatycznej.
2. Model dwuwarstwy lipidowej (1925) - Gortel i Grendel
ekstrahując acetonem lipidy z błon erytrocytów ludzkich i
obliczając powierzchnię błonki utworzonej przez ten ekstrakt,
stwierdzili, że jest ona dwukrotnie większa od powierzchni
wyjściowych krwinek. Sformułowali więc hipotezę, że błona
komórkowa składa się z dwóch warstw lipidowych, sugerując
uwodnienie obu ich stron tzn. polarne główki cząsteczek lipidów
muszą być skierowane na zewnątrz, a niepolarne łańcuchy
węglowodorowe ku sobie, do wnętrza podwójnej warstwy
lipidowej.
3. Model trójwarstwowej błony (1935) - Dowson i Danielli
korzystając z obserwacji Cole, że białka dodane do emulsji
olejowo-wodnej w znacznym stopniu obniżają napięcie
powierzchniowe pomiędzy wodą i kroplami oleju (napięcie takie
jak w naturalnych błonach komórkowych) wysnuli hipotezę, że
błony komórkowe zbudowane są symetrycznie z podwójnej
warstwy lipidowej pokrytej po obu stronach warstwą białek.
4. Model płynnej mozaiki (1972) - Singer i Nicolson opublikowali
teorię modelu płynnej mozaiki w której białka nie tworzą warstwy
na powierzchni lipidów, lecz pływają w dwuwarstwie lipidowej
zanurzone w różnym stopniu. Błona taka jest asymetryczna,
płynna i dynamiczna.
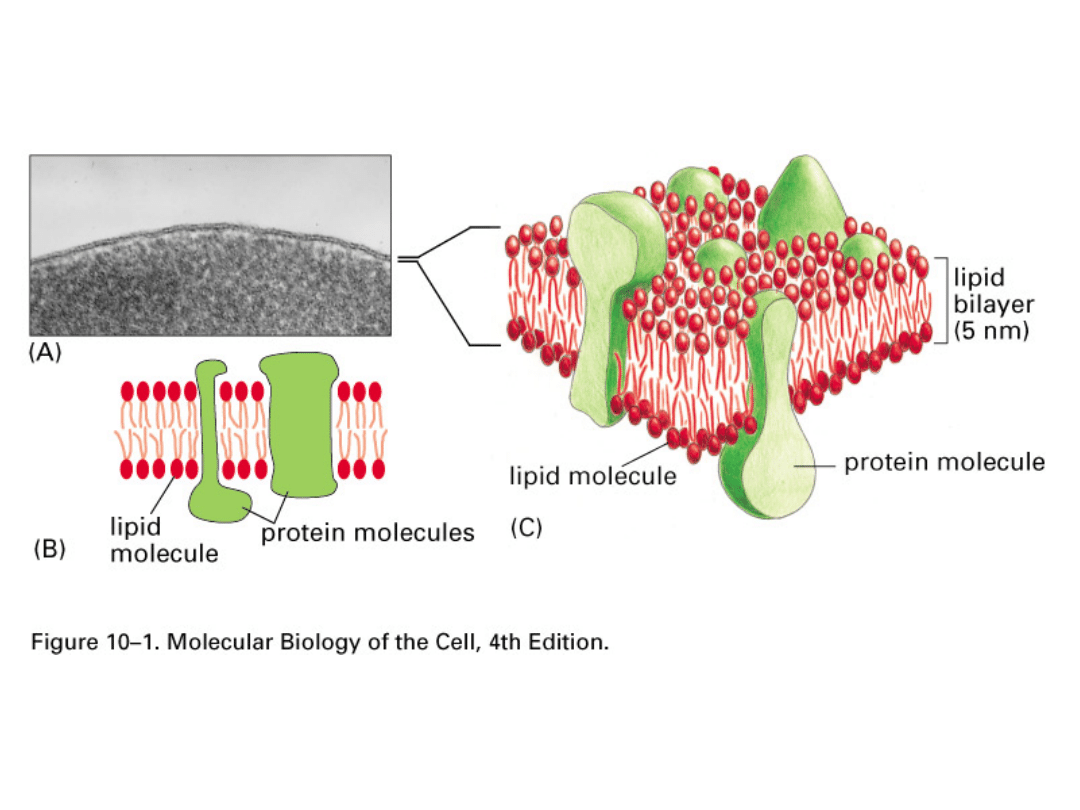

Składniki błon
biologicznych
Wszystkie błony w komórce
zbudowane są z lipidów i białek, oraz
mają wspólny plan budowy ogólnej.
Głównymi składnikami są
lipidy
i
białka
. Wzajemny stosunek tych
składników może być różny w różnych
błonach, a ich ułożenie też bywa
zmienne.
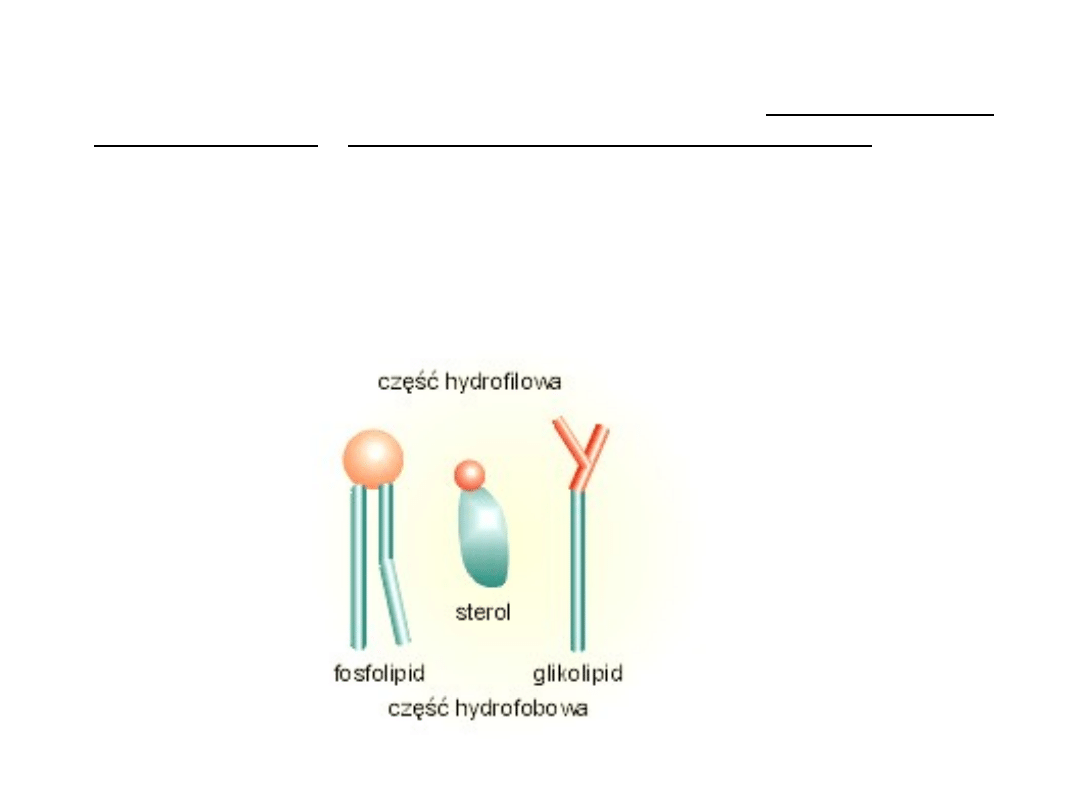
Lipidy w błonach należą do trzech klas: fosfolipidów,
glikolipidów i lipidów obojętnych (sterole).
Podstawową strukturą błony jest dwuwarstwa lipidowa
utworzona z fosfolipidów. Błona taka stanowi ośrodek,
w którym lipidy i białka mogą przemieszczać się po
powierzchni błony a także w poprzek błony.

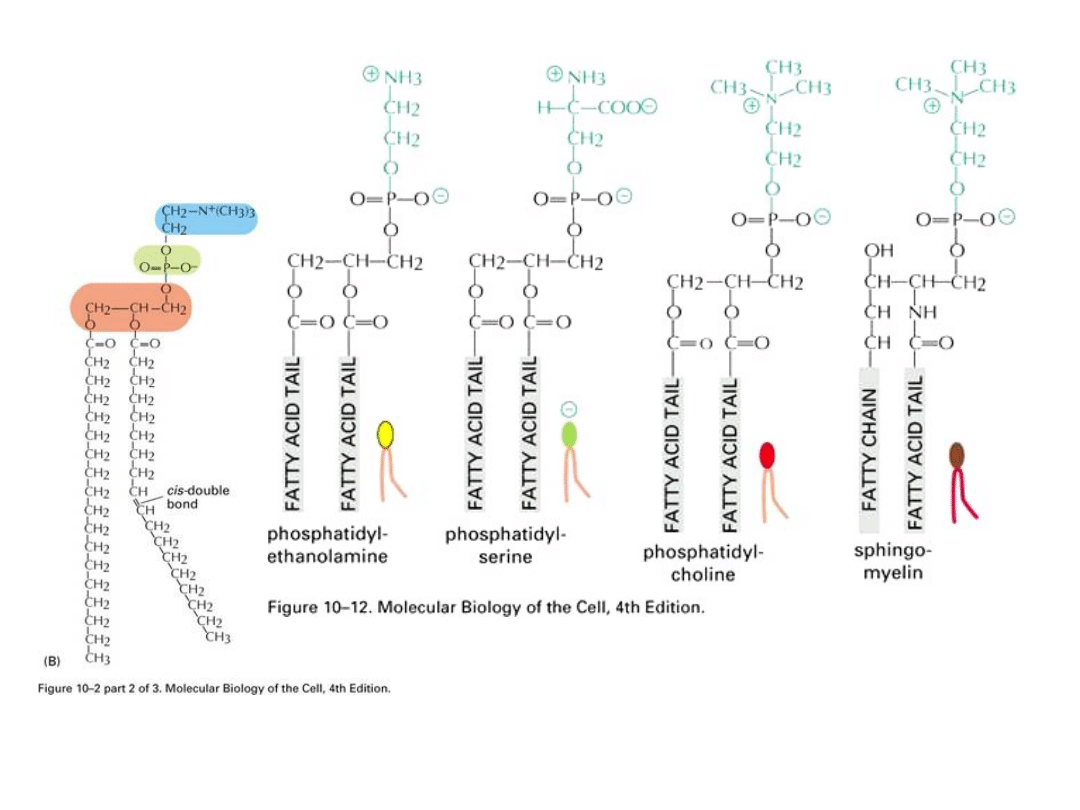
Fosfolipidy
zawierają dwie cząsteczki kwasów
tłuszczowych połączone z dwoma spośród trzech atomów
węgla glicerolu. Trzeci węgiel w glicerolu połączony jest z
ujemnie naładowaną hydrofilową grupą fosforanową do której z
kolei jest przyłączony mały związek hydrofilowy, taki jak
cholina. Każda cząsteczka fosfolipidu zawiera więc
hydrofobowy „ogon", złożony z dwóch łańcuchów kwasu
tłuszczowego, oraz hydrofilową „głowę", gdzie znajduje się
fosforan. Cząsteczki takie jak fosfolipidy, z regionami zarówno
hydrofobowymi jak i hydrofilowymi, są nazywane cząsteczkami
amfipatycznymi.
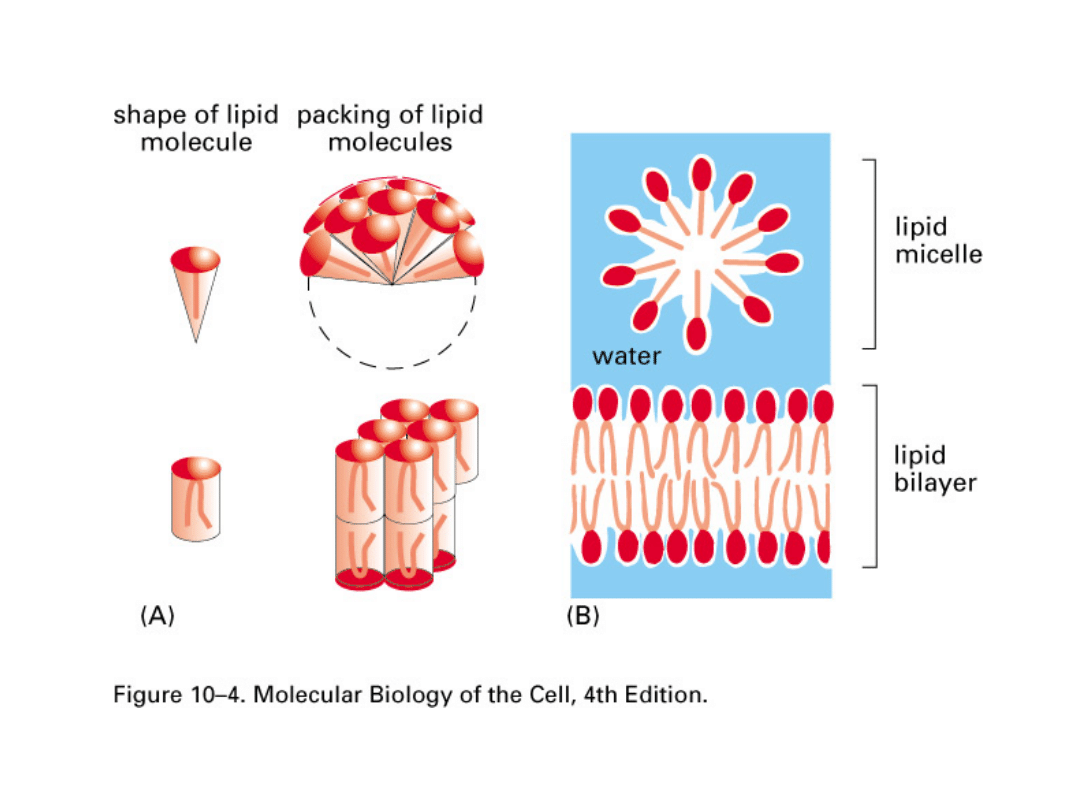
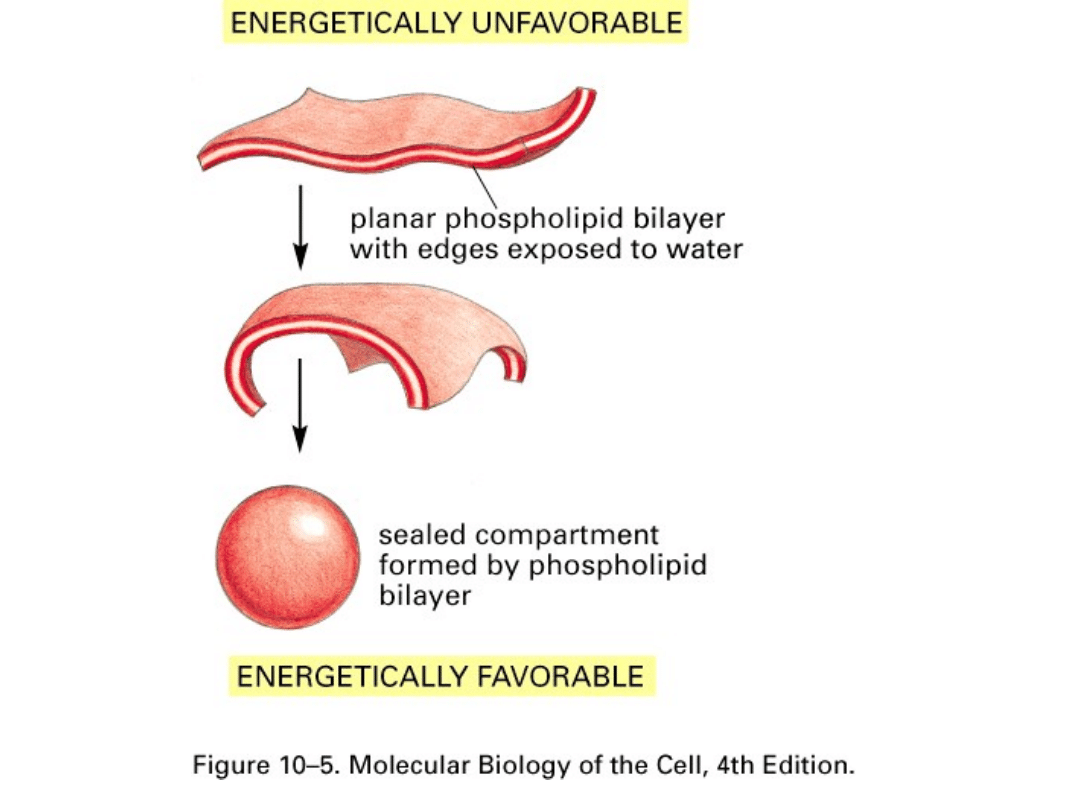
Zdolność fosfolipidów do tworzenia błon jest związana z ich
amfipatycznym charakterem. Fosfolipidy rozprzestrzeniają się
na powierzchni wody, tworząc pojedynczą warstwę cząsteczek
fosfolipidowych, z hydrofobowymi „ogonami" skierowanymi ku
górze, i hydrofilowymi „głowami" kontaktującymi się z wodą.
Dwie takie jednocząsteczkowe warstwy mogą łączyć się na
zasadzie „ogon z ogonem", tworząc dwuwarstwę
fosfolipidową. Taka orientacja jest najbardziej korzystna pod
względem energetycznym, gdyż pozwala na swobodny kontakt
hydrofilowych głów z wodą, podczas gdy hydrofobowe łańcuchy
kwasów tłuszczowych unikają kontaktu z wodą, gromadząc się
w środku układu. Dodatkowo cząsteczki fosfolipidów mają w
przybliżeniu jednakową szerokość, co również sprzyja układaniu
się ich w podwójne warstwy cylindrycznych struktur.

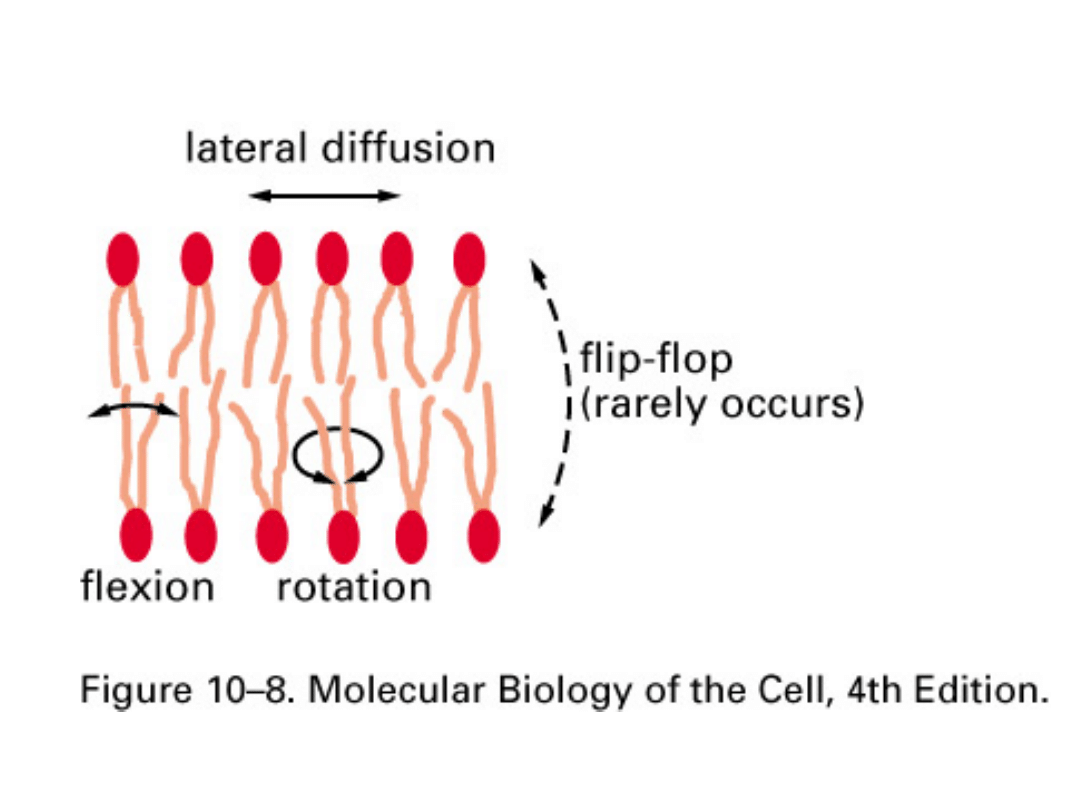
Cząsteczka fosfolipidu w błonie nie jest sztywna. Oprócz
ruchów obrotowych całej cząsteczki wokół swojej osi
występuje rozchodzenie się i zginanie łańcuchów kwasów
tłuszczowych. Mniej ruchliwa jest okolica polarna cząsteczki,
natomiast schowane w głębi warstwy hydrofobowej końce
łańcuchów węglowodorowych wykonują szybkie ruchy.
Ruchliwość łańcucha węglowodorowego jest tym większa im jest
on krótszy i ma liczniejsze wiązania nienasycone. Fosfolipidy
łatwo przemieszczają się w obrębie jednej warstwy
lipidowej błony (dyfuzja boczna) - zachodzi co około 10
-6
sekundy. Natomiast wymiana cząsteczek lipidów między
jedną i drugą warstwą (tzw. ruchy flip-flop) może być
bardzo wolna i zachodzić raz na kilkaset godzin.
W komórkach bakterii i drożdży, które muszą adaptować się do
różnych temperatur, zarówno długość jak i stopień nienasycenia
kwasów tłuszczowych są stale dopasowywane, tak aby
utrzymać względnie stały poziom płynności błony: w wyższych
temperaturach komórka wytwarza lipidy o łańcuchach dłuższych
i zawierających mniej wiązań podwójnych, co sprzyja
zachowaniu stabilności i płynności błony.
Płynność błon umożliwia fuzję błon ze sobą i mieszanie się ich
składników, co przy podziale komórki zapewnia równomierne
rozdzielenie budujących błonę cząsteczek pomiędzy komórki
potomne.

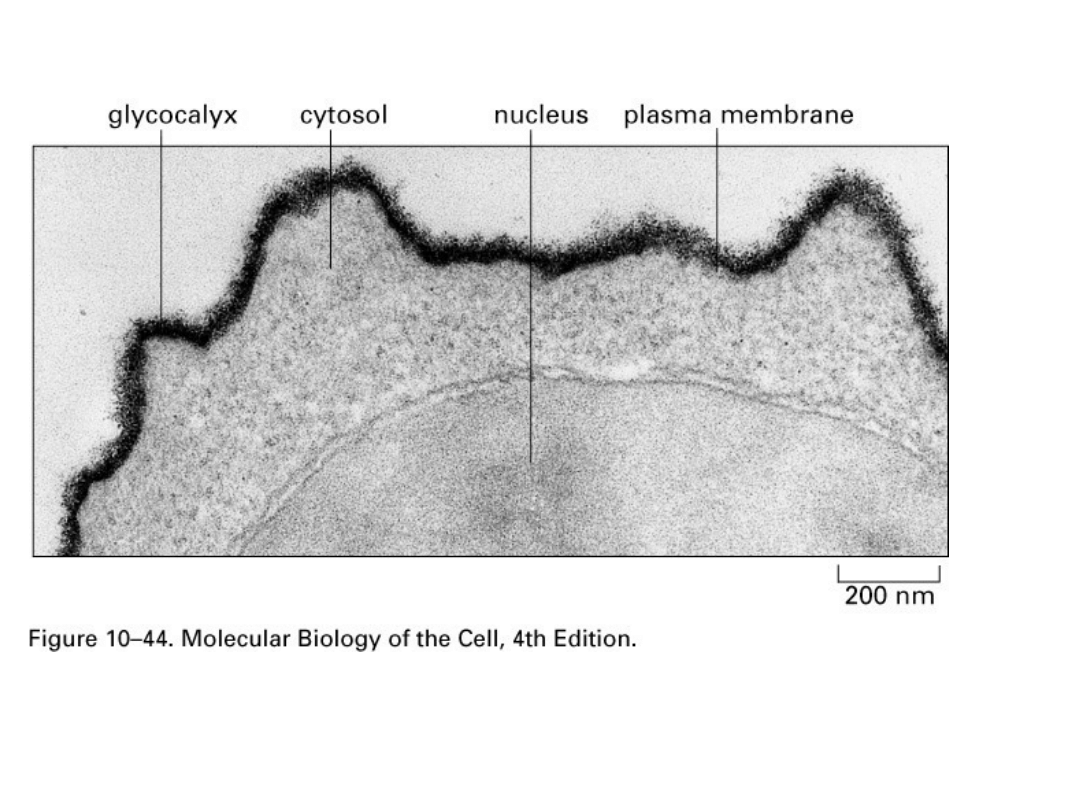
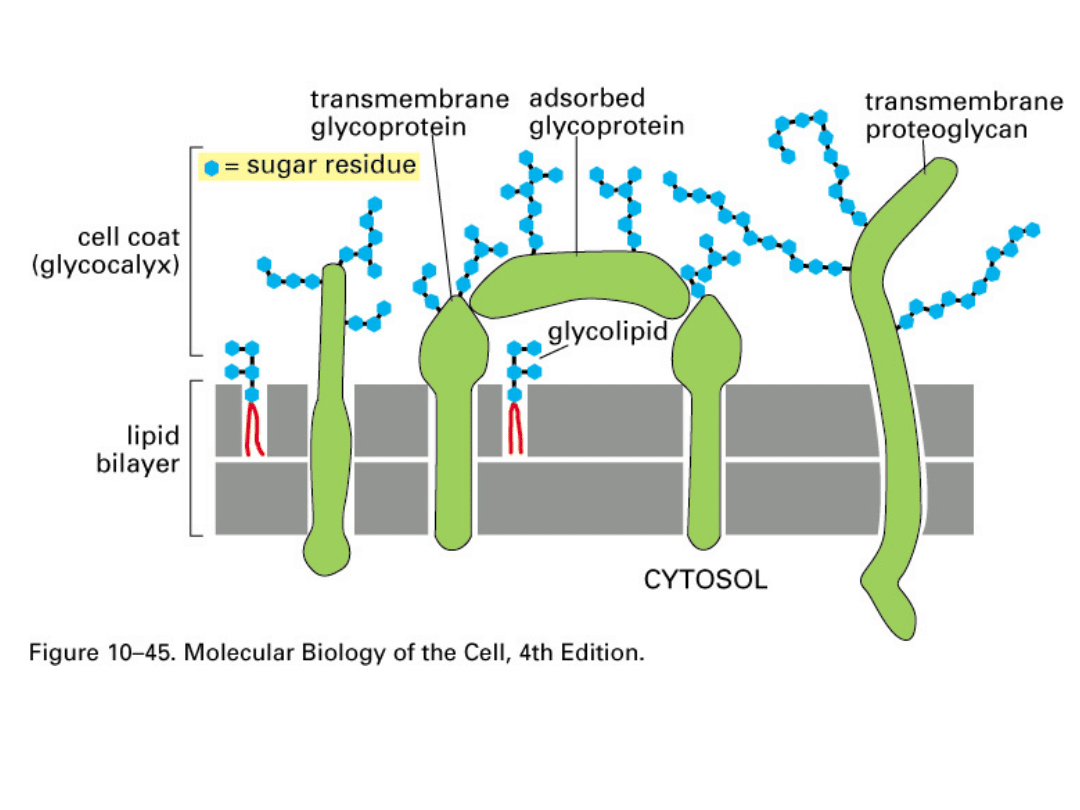
Glikolipidy - są to cząsteczki lipidów połączone z
łańcuchami polisacharydowymi. Zlokalizowane są w
zewnętrznej warstwie błony. Domeny polarne glikolipidów
wystają ponad powierzchnię błony komórkowej, prezentując
swoje grupy polarne do środowiska.
Jakkolwiek rola glikolipidów nie jest do końca poznana, to
przypisuje się im rozmaite funkcje:
1) utrzymują asymetryczność błony komórkowej,
2) oddzielają komórki od środowiska i stabilizują błonę
komórkową,
3) są receptorami dla niektórych hormonów peptydowych i
toksyn bakteryjnych,
4) dzięki specyficznej kombinacji topograficznej reszt cukrowych
w błonach erytrocytów określają grupy krwi (ABO).
Glikolipidy są na tyle ważnymi składnikami błon, że w
przypadku wad genetycznych związanych z ich
metabolizmem występują duże zaburzenia rozwojowe,
kończące się przedwczesną śmiercią noworodka.
Warstwa glikolipidów pokrywa większość komórek
zwierzęcych tworząc tzw. glikokaliks. Glikolipidy uzyskują
swoje grupy cukrowe w aparacie Golgiego.

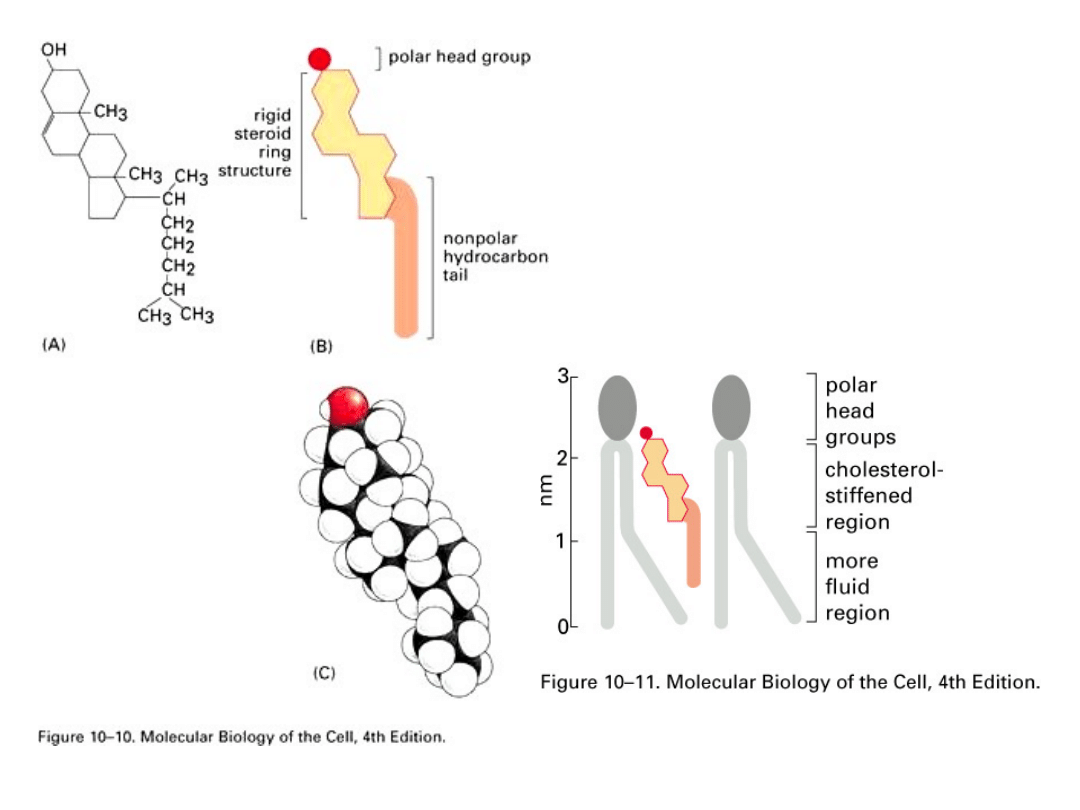
Sterole - zbudowane są ze sztywnego poczwórnego pierścienia
węglowego z bocznymi podstawnikami. W komórkach zwierzęcych
głównym sterolem (steroidem) jest cholesterol, zaś u roślin
występują fitosterole: sitosterol, kamposterol i stigmosterol. W
błonie lokalizują się pomiędzy łańcuchami węglowodorowymi
fosfolipidów. Cholesterol jest lipidem o słabych właściwośćiach
amfipatycznych. Jego cząsteczka składa się z części hydrofobowej
– steroidowej i łańcucha alifatycznego dołączonego do węgla 17 w
pierścieniu D. Domena hydrofilowa reprezentowana jest przez
grupę (OH-), związaną z 3 węglem w pierścieniu A. Cholesterol jest
umiejscowiony w błonie komórkowej, podobnie jak glikolipidy, w jej
zewnętrznej warstwie. W niej wiąże się swoją grupą hydroksylową
z 1 węglem łańcucha alifatycznego kwasu tłuszczowego
fosfolipidu. Cholesterol jest podstawowym czynnikiem
regulującym przepuszczalność błon komórkowych.
Położenie grupy hydrofobowej pomiędzy łańcuchami
alifatycznymi fosfolipidów zapobiega przejściu fazowemu
dużych obszarów błony (zapobiega zbytniemu zbliżaniu się
łańcuchów i uniemożliwia powstawanie pomiędzy nimi
oddziaływań van der Waalsa, co prowadziło by do ich
unieruchomienia i przejście w stan stały), utrzymuje
wewnętrzną, hydrofobową część dwuwarstwy lipidowej w
stanie płynnym. Natomiast grupy polarne cholesterolu
uszczelniają oraz usztywniają i stabilizują zewnętrzne
krawędzie dwuwarstwy lipidowej, zapobiegając
niekontrolowanej migracji małych cząstek rozpuszczalnych
w wodzie pomiędzy cząsteczkami fosfolipidów.

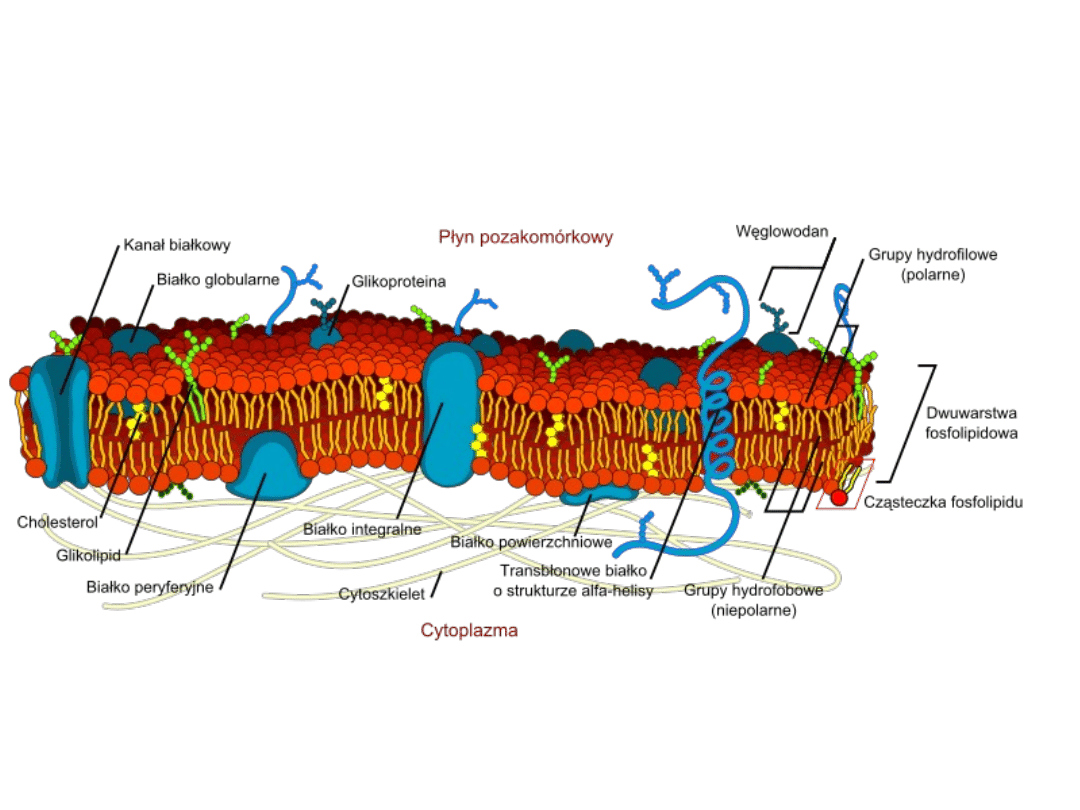
Białka błonowe
umownie dzieli się
na dwie grupy:
• powierzchniowe (peryferyjne)
• integralne

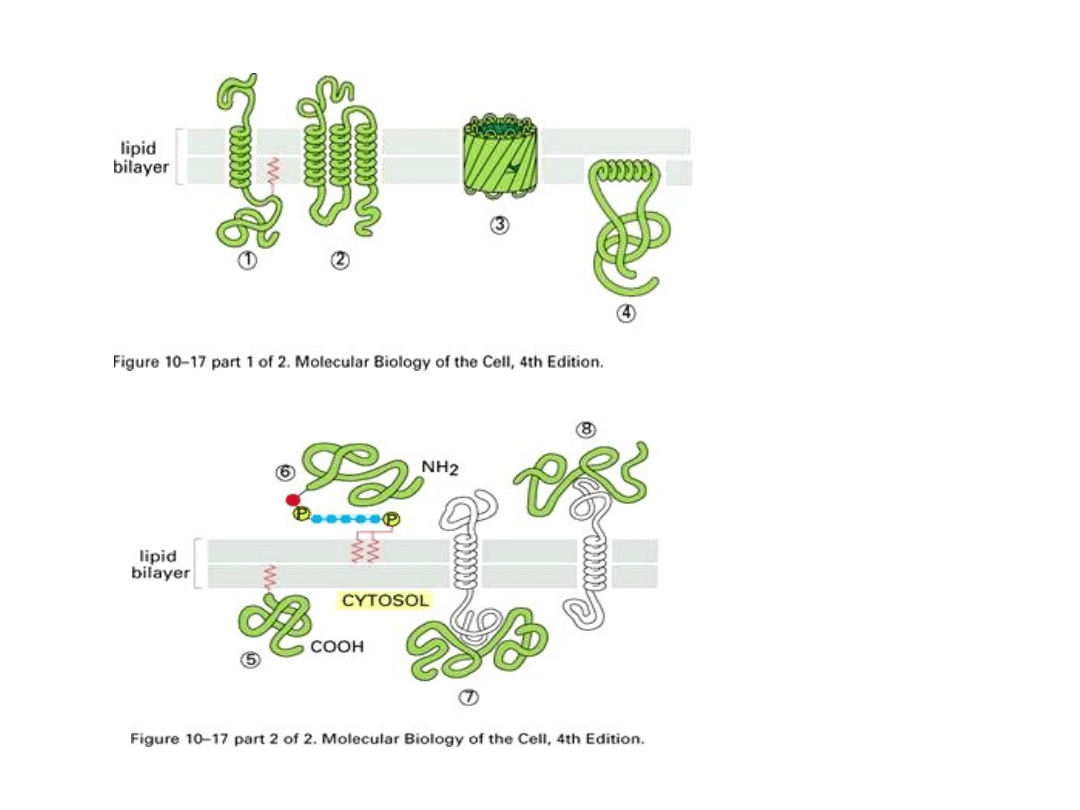
Schemat organizacji białek błonowych
1. Białko transmembranowe
2. Białko monowarstwy zewnętrznej
3. Białko monowarstwy wewnętrznej
4. Białko wewnętrzne błony
Niebieskie – białka peryferyjne

• Białka które dają się łatwo usunąć z błony wodą,
roztworami soli lub czynników chelatujących nie
niszcząc dwuwarstwy lipidowej - są to
BIAŁKA
POWIERZCHNIOWE
(peryferyjne) błony. Są one
luźno związane z powierzchniami błony i często
połączone z łańcuchami sacharydowymi (glikoproteiny)
oraz kwasami tłuszczowymi czy długołańcuchowymi
alkoholami, poprzez które polipeptydy te zakotwiczają
się w obrębie błony. Białka powierzchniowe są
cząsteczkami hydrofilnymi i najczęściej występują w
rejonach,
w których sterczą z błon fragmenty białek integralnych
i są z nimi powiązane oddziaływaniami
niekowalencyjnymi. Mogą również wiązać się
z polarnymi fragmentami fosfolipidów. Część białek
może znajdować się całkowicie poza rejonem błony,
a jedynie wiązać się z nią za pomocą kowalencyjnego
wiązania z cząsteczką lipidową błony.

• Te które można wyizolować z błony
do roztworu wodnego jedynie w
postaci kompleksów z detergentem
(solubilizacja detergentem -
przeprowadzenie do roztworu
wodnego kompleksów detergentu i
składników błony) niszczącym
uporządkowanie dwuwarstwy
lipidowej - są to
BIAŁKA
INTEGRALNE
- na trwałe
wbudowane w dwuwarstwę

• Białka integralne mogą być zbudowane z jednej lub kilku
podjednostek. Fragmenty cząsteczek białkowych mogą
wyłaniać się na jednej lub na obu powierzchniach błony,
bądź są prawie całkowicie schowane w części hydrofobowej
dwuwarstwy lipidowej. Białka integralne mają w łańcuchu
polipeptydowym przynajmniej jedną sekwencję składającą
się z co najmniej 22 aminokwasów hydrofobowych, które
pozwalają na zakotwiczenie się w błonie. W niektórych
białkach reszty aminokwasów hydrofobowych tworzą kilka
skupień, co sprawia, że łańcuch polipeptydowy kilkakrotnie
przemierza dwuwarstwę lipidową. Koniec karboksylowy [C]
łańcuchów polipeptydowych czasem jest skierowany do
cytoplazmy, a koniec aminowy [N] na powierzchnię
zewnętrzną błony, może też być przeciwnie. Białka mogą
również kotwiczyć się w błonie poprzez kowalencyjnie
związane z nimi łańcuchy kwasów tłuszczowych lub
cząsteczkę glikofosfolipidu. Białka błonowe rozmieszczone
są w błonie asymetrycznie. Ich ułożenie nie jest
przypadkowe, ale wynika ze specyficznych oddziaływań
łańcucha polipeptydowego z dwuwarstwą lipidową.
Wszystkie te cechy białek integralnych przyczyniają się do
asymetrii błony. Większość białek integralnych błon
biologicznych jest glikoproteinami.
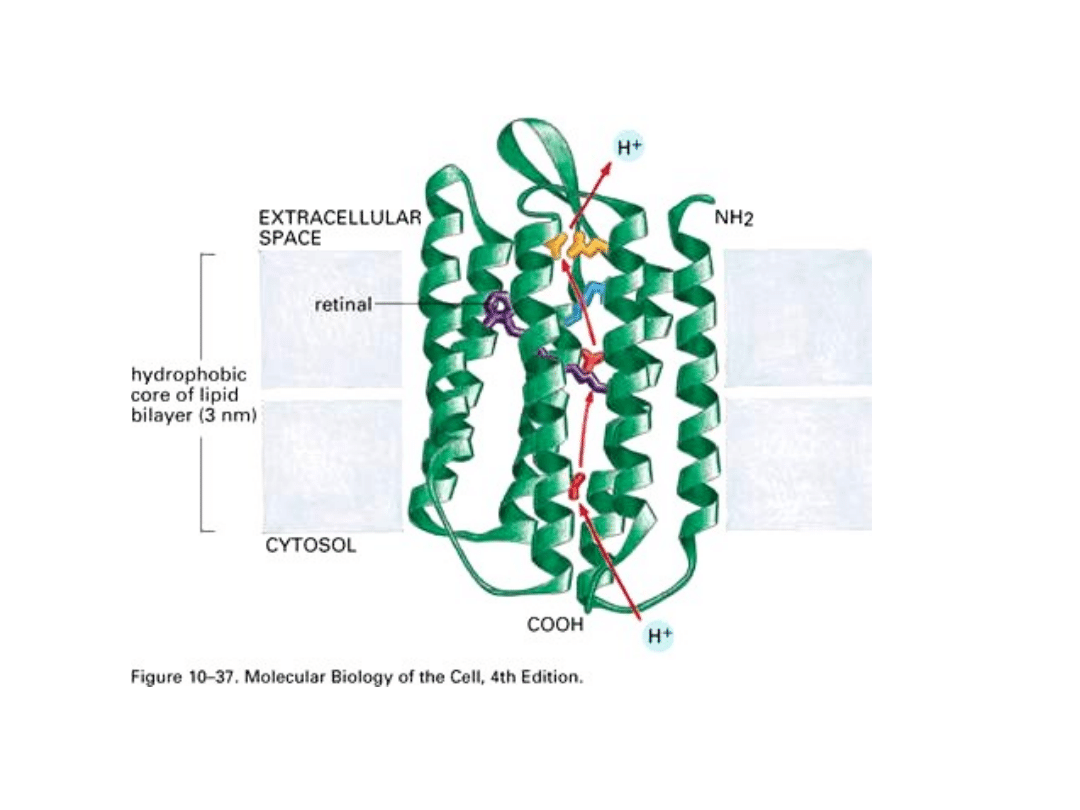

Funkcje białek błonowych
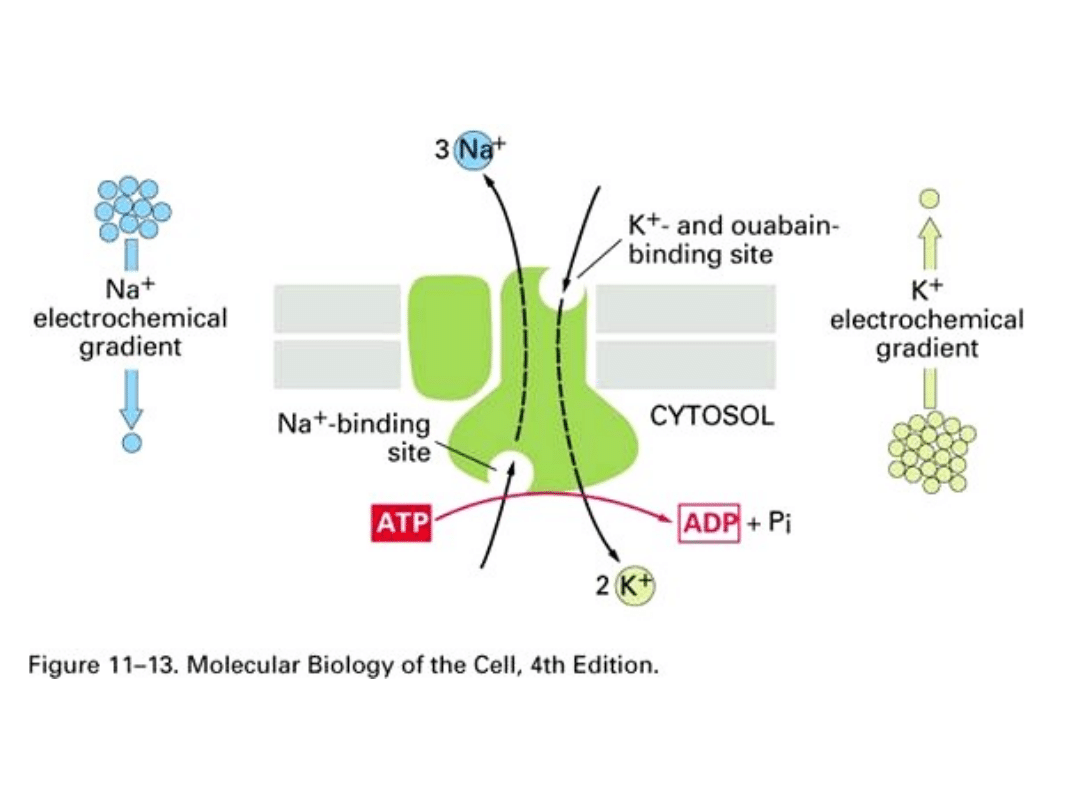
• Białka transportujące
– uczestniczą w transporcie przez błony małych
cząsteczek, tworzą kanały i pompy prowadząc transport kontrolowany (np.
pompa sodowa, aktywnie wypompowuje z komórki jony sodu i wprowadza do niej
jony potasu).
• Białka wiążące
– są elementami wyspecjalizowanych struktur odpowiedzialnych
za utrzymywanie łączności pomiędzy komórkami lub z cytoszkieletem (np.
integryny wiążące elementy wewnątrzkomórkowe filamenty aktyny z białkami
substancji zewnątrzkomórkowej).
• Białka receptorowe
– pośredniczą w przekazywaniu informacji ze środowiska
zewnętrznego do komórki, związanie cząsteczki sygnałowej indukuje zmiany w
aktywności komórkowej (np. receptor płytkopochodnego czynnika wzrostu, który
wytwarza wewnątrzkomórkowe sygnały powodujące wzrost i podział komórki).
• Białka enzymatyczne
– enzymy, których miejsca katalityczne znajdują się po
jednej ze stron błony bądź w jej wnętrzu (np. cyklaza adenylanowa, w odpowiedzi
na sygnały zewnątrzkomórkowe katalizuje wytwarzanie wewnątrzkomórkowego
cyklicznego AMP, będącego wewnątrzkomórkowym przekaźnikiem informacji)
Uważa się, że białka integralne pełniące funkcje transportowe, których łańcuch
polipeptydowy wielokrotnie przemierza dwuwarstwę lipidową - tworzy przez
błonę kanały. Modele kanałów błonowych przyjmują, że 22-aminokwasowe
hydrofobowe odcinki łańcucha polipeptydowego tworzą struktury a-helisy, a kilka
takich a- helis obok siebie stanowi ścianę kanału. Oprócz tych zewnętrznych a-
helis mocujących kanał w dwuwarstwie lipidowej, wewnątrz kanału mogą biec
dodatkowe, wewnętrzne odcinki łańcucha, zbudowane z hydrofilnych
aminokwasów. Pełnią one właściwe funkcje transportowe np. białko kanałów
wapniowych.

WŁAŚCIWOŚCI BŁON BIOLOGICZNYCH
• Półpłynność:
dwuwarstwa lipidowa błony biologicznej jest
w stanie półpłynnym lub inaczej płynno-krystalicznym. Ze
względu na wysoki stopień uporządkowania ma ona
właściwości krystaliczne (fosfolipidy ułożone w szeregi,
biegunem polarnym na zewnątrz, apolarnym do środka).
Z drugiej zaś strony podwójna warstwa lipidowa wykazuje
właściwości płynne, bowiem pomimo tego uporządkowania
łańcuchy węglowodorowe pozostają w ciągłym ruchu, co
oznacza, że cząsteczki fosfolipidów mają swobodę rotacji
i mogą dyfundować w obrębie pojedynczej warstwy błony,
w której występują. Nadaje to podwójnej warstwie
fosfolipidowej charakter cieczy krystalicznej, który bywa
też określany jako półpłynny. Niektóre błony biologiczne
w temperaturze optymalnej dla wzrostu komórki zawierają
jednak pewne lipidy w formie krystalicznej. Krystaliczna
struktura jest stanem, w którym cząsteczki lipidów są
względem siebie uporządkowane, co powoduje ich
wzajemne powiązanie a tym samym unieruchomienie .

WŁAŚCIWOŚCI BŁON BIOLOGICZNYCH
• Dynamiczność:
jest wyrażona w ruchach budujących błonę
lipidów i białek. Cząsteczki fosfolipidów w błonie nie są sztywne. Mniej
ruchliwe są ich okolice polarne, natomiast zanurzone w głębi warstwy
hydrofobowej końce łańcuchów węglowodorowych wykonują szybkie
ruchy, tym szybsze im te łańcuchy są krótsze i zawierają liczniejsze
wiązania podwójne. Białka błony mogą natomiast być w jej płaszczyźnie
przemieszczane dyfuzyjnie, wykonywać ruchy obrotowe w osi
prostopadłej do powierzchni błony oraz wynurzać się z dwuwarstwy
lipidowej lub w niej zanurzać.
Ruchliwość składników błon powoduje zamykanie wszelkich wyrw i
ubytków. Błony w żywych komórkach nigdy nie tworzą wolnych krawędzi.
Dzięki temu wnętrze komórki i poszczególnych jej przedziałów jest
zawsze otoczone selektywnie przepuszczającą barierą. Ponieważ błona
jest dwuwymiarowym płynem, wiele jej białek, podobnie jak i lipidów,
może swobodnie poruszać się w obrębie płaszczyzny dwuwarstwy
lipidowej. Można to w sposób łatwy i oczywisty wykazać, doprowadzając
do fuzji komórki myszy z komórką ludzką, tworząc podwójnej wielkości
komórkę hybrydową, a następnie śledząc rozmieszczenie białek błony
komórkowej zarówno myszy, jak i człowieka. Aczkolwiek na początku
białka te pozostaną na powierzchni swych odpowiednich połówek nowo
powstałej komórki hybrydowej, to już po niecałej godzinie dwa zestawy
tych białek zostaną równomiernie wymieszane na całej powierzchni
komórki.

WŁAŚCIWOŚCI BŁON BIOLOGICZNYCH
• Asymetryczność:
polega na różnicach w budowie obu
powierzchni błony, skierowanych na zewnątrz i ku wnętrzu
komórki lub organelli. Dwie warstwy dwuwarstwy często
zawierają różny skład fosfolipidów i glikolipidów a białka są
wtopione w dwuwarstwę ze specyficzną orientacją przestrzenną,
konieczną dla ich funkcji.
W błonie komórkowej (plazmolemie) wyróżnia się dwie
warstwy:
– warstwę lipidową cytoplazmatyczną P (ang. protoplasmic)
od strony protoplazmy
– warstwę lipidową zewnętrzną E (ang. exoplasmic) od strony
środowiska.
Asymetria dwuwarstwy lipidowej błony komórkowej jest
utrzymywana głównie przez obecność glikolipidów i
glikosacharydów, które wchodzą w skład zewnętrznej (E)
warstwy błony, a ich reszty cukrowe są eksponowane na
zewnątrz komórki. Przykładem glikolipidów mogą być cząsteczki
noszące własności grupowe ABO erytrocytów człowieka.

WŁAŚCIWOŚCI BŁON BIOLOGICZNYCH
• Półprzepuszczalność (selektywność):
przez błonę mogą swobodnie
przenikać tylko nieliczne związki np. H
2
O, CO
2
, glicerol; natomiast
większość substancji, aby mogła przeniknąć przez błonę wymaga
obecności w błonie odpowiednich układów transportujących, którymi
są odpowiednie białka błonowe. Przepuszczalność błony dla danej
substancji zależy od rozmiaru
i ładunku jej cząsteczki. Na przykład cząsteczki wody z dużą szybkością
przedostają się przez szczelinę w podwójnej warstwie lipidowej,
powstałą na skutek chwilowego odchylenia się łańcucha kwasu
tłuszczowego. Bez trudu przez dwuwarstwę przenikają gazy, np. tlen,
CO
2
i N
2
, małe cząsteczki polarne, np. glicerol, i niektóre większe
cząsteczki apolarne (hydrofobowe), np. węglowodory. Cząsteczki
większe, np. glukoza i jony różnej wielkości nie przedostają się z
powodu zbyt dużych rozmiarów lub na skutek odpychania przez
ujemnie naładowaną powierzchnię błony. Przepuszczalność dla tych
związków wiąże się z występowaniem w błonie specyficznych białek
transportujących. Wszystkie błony plazmatyczne są selektywnie
przepuszczalne dla różnych rodzajów cząsteczek a wynika to z
występowania w poszczególnych typach błon różnych zestawów białek
transportujących.
W odpowiedzi na zmianę warunków środowiska lub na aktualne
zapotrzebowanie komórki błona może czasami stawać się barierą dla
danej substancji, w innych natomiast okolicznościach może je aktywnie
transportować. Kierując ruchem cząsteczek, komórka jest w stanie
zapewnić stałość składu jonowego i cząsteczkowego swego
wewnętrznego środowiska.
Schemat membrany półprzepuszczalnej
podczas hemodializy

WŁAŚCIWOŚCI BŁON BIOLOGICZNYCH
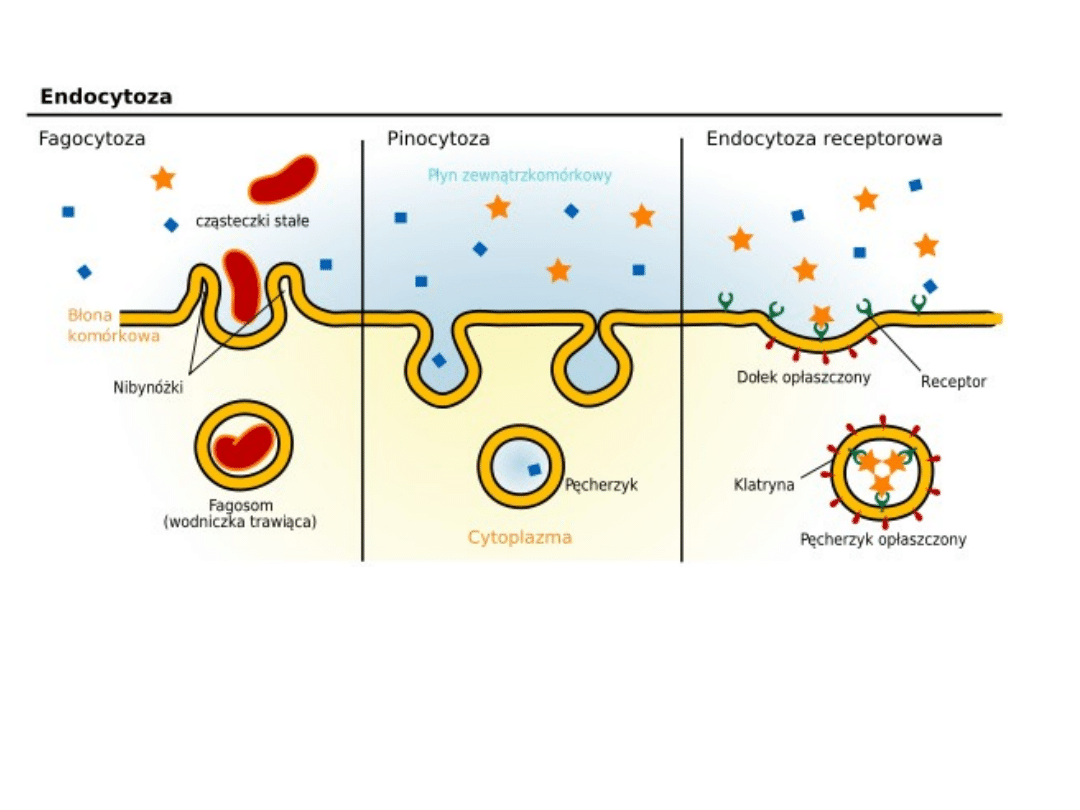
• Zdolność do fuzji:
ważną cechą podwójnych warstw
lipidowych jest unikanie tworzenia układów z wolnymi
końcami, czego wyrazem jest spontaniczne zamykanie się
błon w struktury pęcherzykowate, oraz zdolność w
określonych warunkach do łączenia się z innymi,
podobnymi strukturami błonowymi. Fuzja (łączenie się,
zlewanie) błon jest powszechnie występującym
procesem i ma istotne znaczenie dla funkcjonowania
komórki, np. podczas endocytozy, wydzielania i
krążenia składników błon. Zachodzi też w
wyspecjalizowanych komórkach, np. podczas
egzocytozy (wydzielania) enzymów, neurohormonów,
podczas łączenia się komórki jajowej z plemnikiem,
łączenia się mioblastów. Fuzja zachodzi też w procesach
patologicznych, np. w odpowiedzi zapalnej podczas
tworzenia się komórek olbrzymich, podczas wnikania do
komórki wirusów z otoczką.
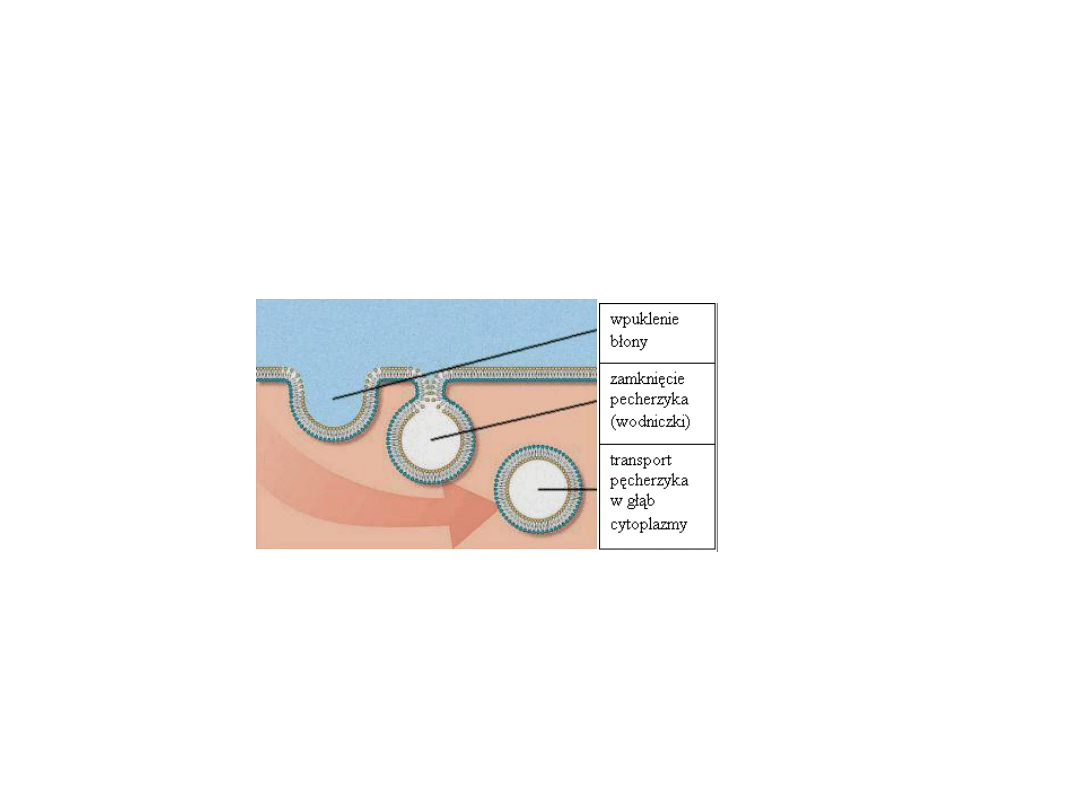
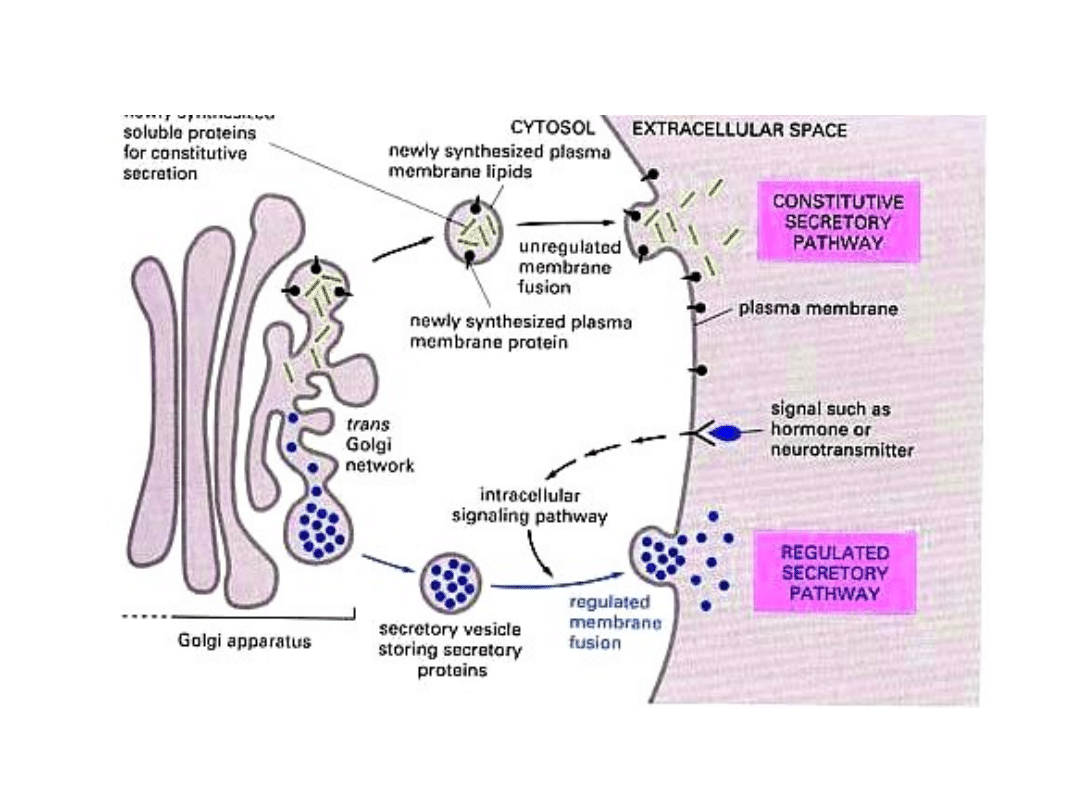
przebieg endocytozy
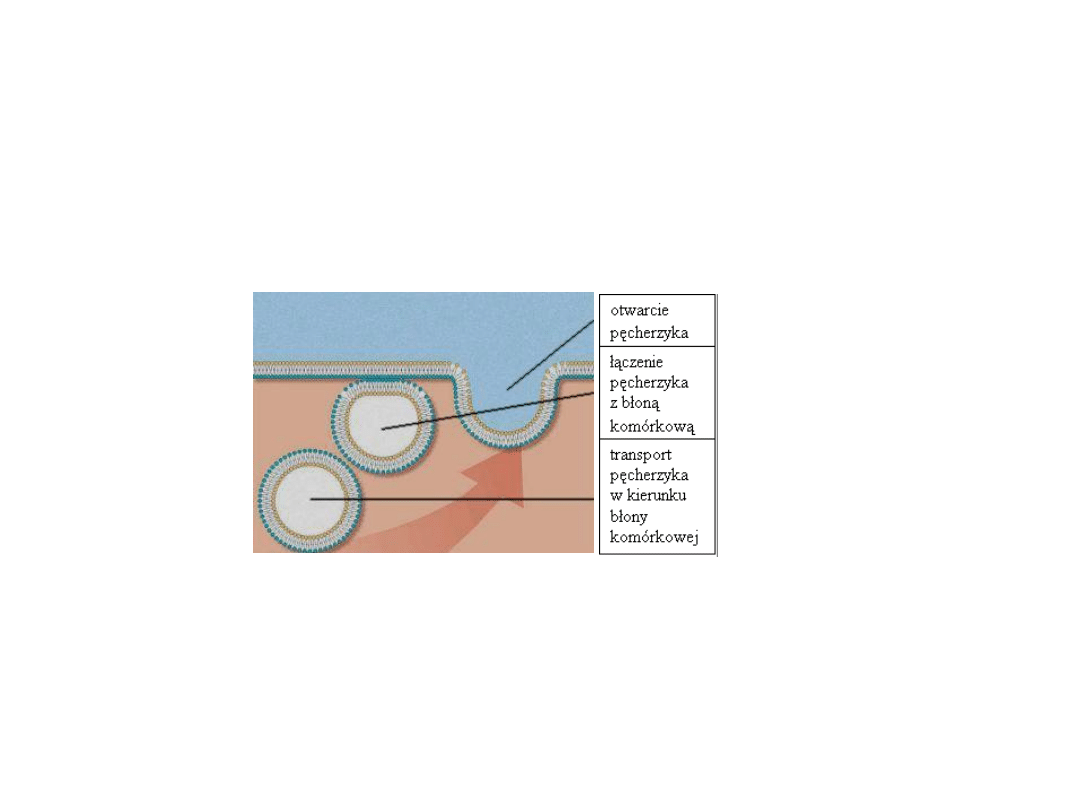
przebieg egzocytozy

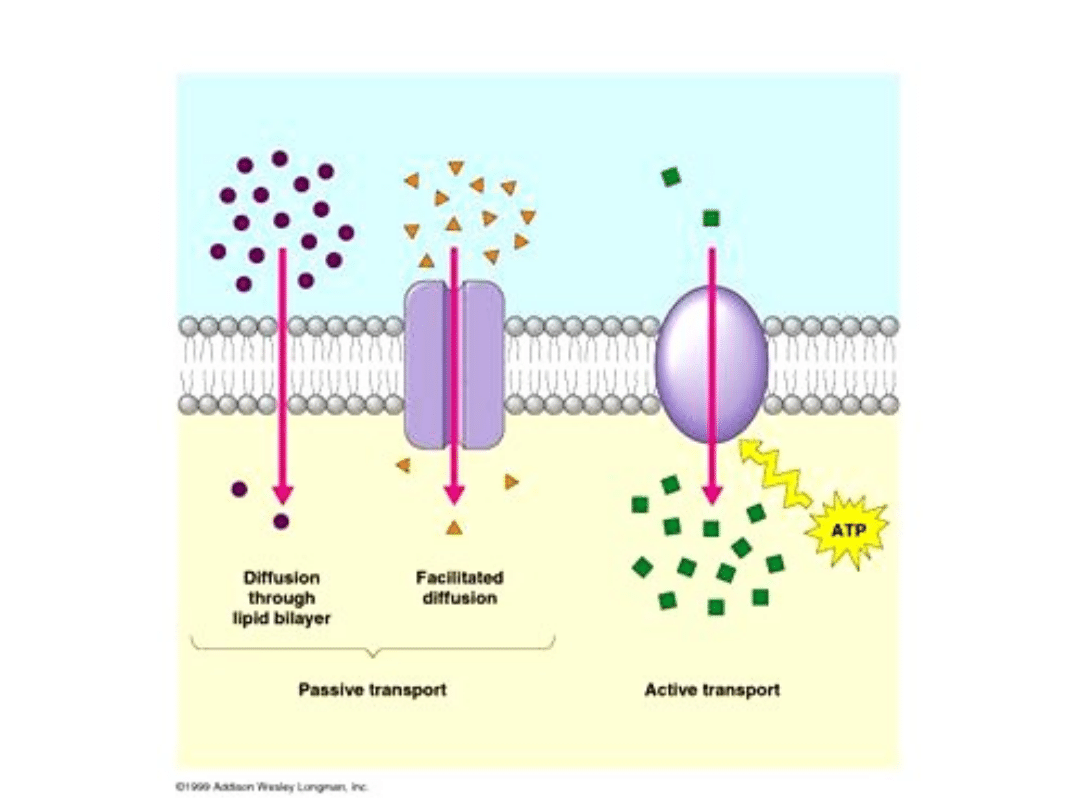
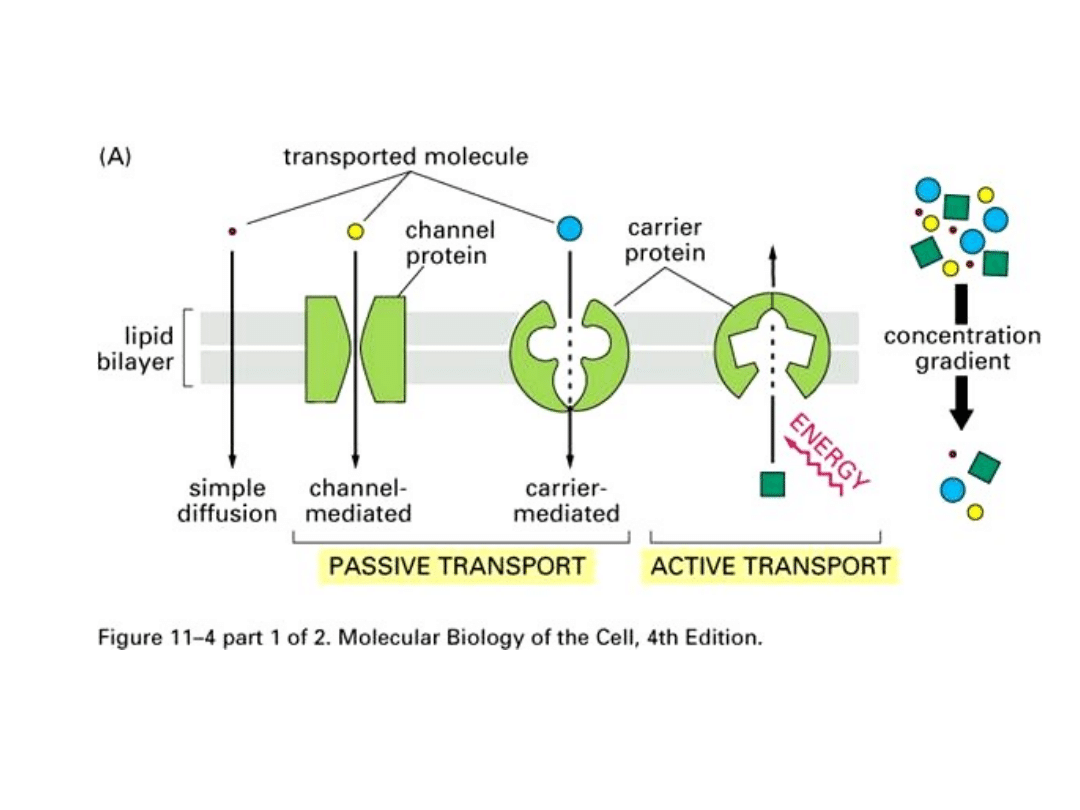
Transport bierny
Dyfuzja prosta to samorzutny ruch cząsteczek danej substancji
rozpuszczonej od stężenia większego do mniejszego (zgodnie z
gradientem stężeń) Im wyższa temperatura tym szybciej
zachodzić będzie dyfuzja.
Osmoza to szczególny przypadek dyfuzji - mianowicie dyfuzja
przez błonę półprzepuszczalną (nie przepuszczająca substancji
rozpuszczonych). W komórce takimi błonami, przez które zachodzi
osmoza są plazmalema (błona komórkowa) i tonoplast (błona
wakuoli). Ciśnienie osmotyczne jest wprost proporcjonalne do
temperatury i do stężenia roztworu.
Dyfuzja wspomagana (ułatwiona) nie wymaga zużycia energii
z ATP, aczkolwiek wymaga specyficznych nośników (białek
błonowych, kanałów białkowych itp.) które umożliwiają substancji
rozpuszczonej przeniknięcie przez normalnie nieprzepuszczalną
błonę. Dyfuzja wspomagana odbywa się jak każda dyfuzja,
zgodnie z gradientem stężeń. Przykładem może być wchłanianie
fruktozy w jelicie cienkim.

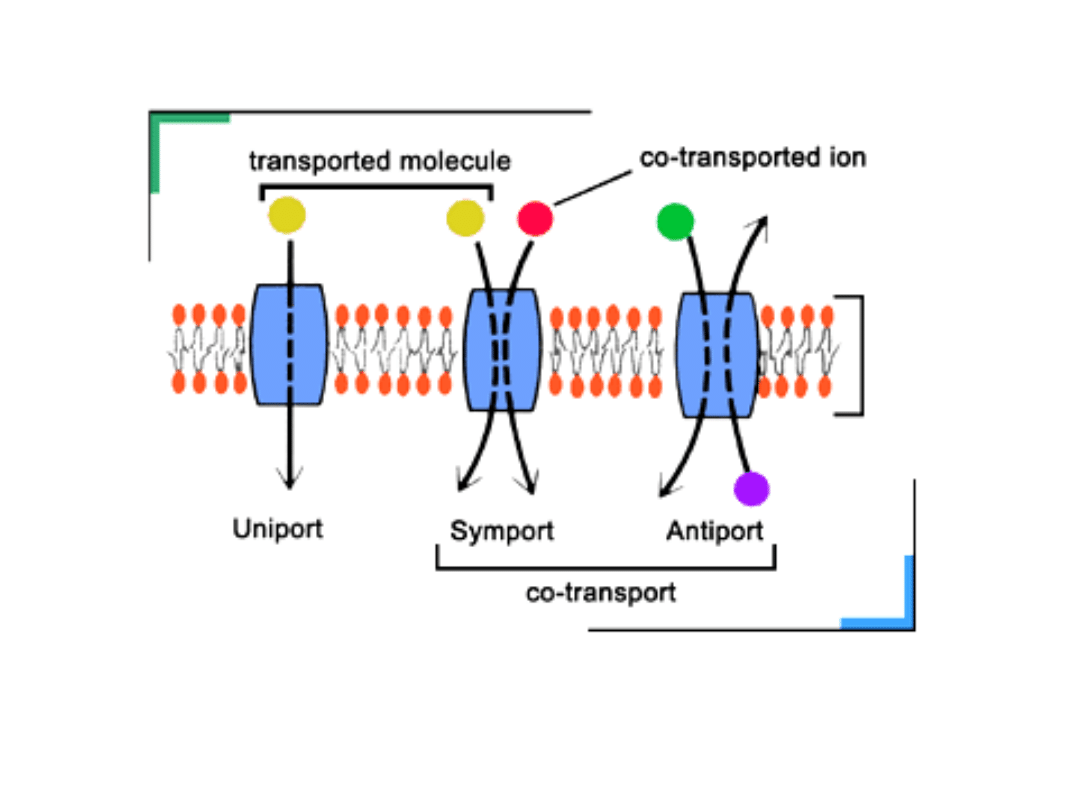
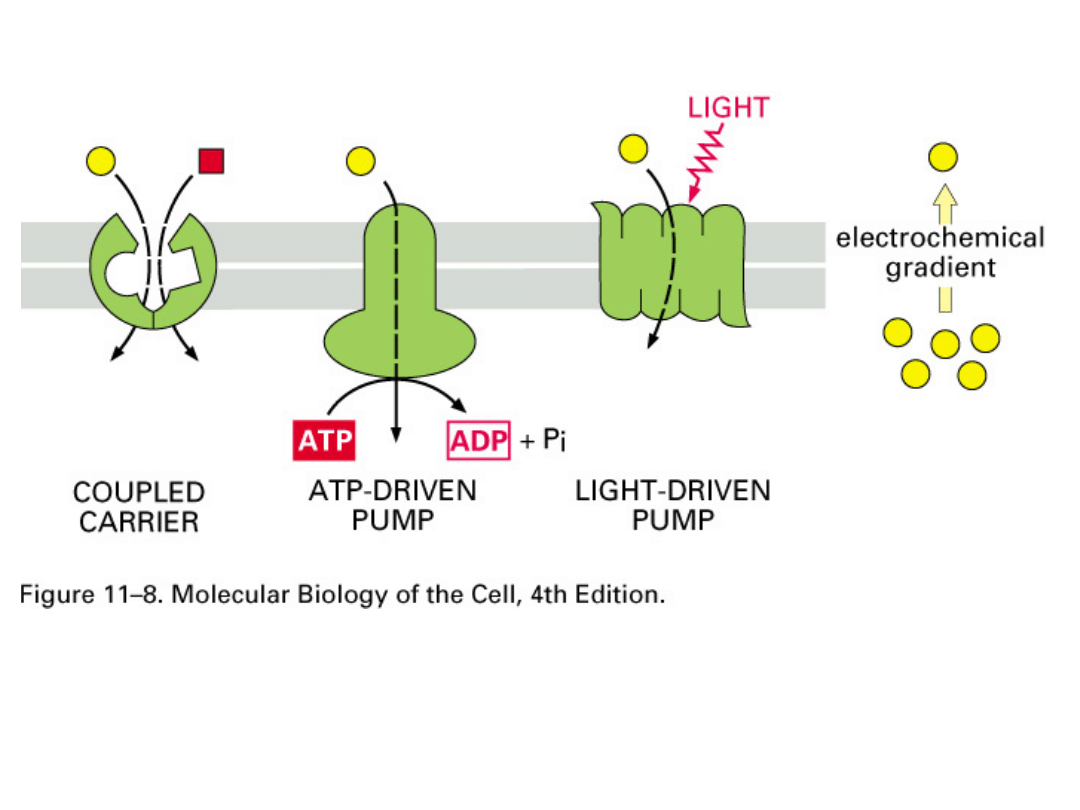
Transport aktywny
- rodzaj przenikania związków
chemicznych przez błony biologiczne, który zachodzi
z udziałem pewnych mechanizmów transportujących
lub substancji przenośnikowych, ze środowiska o
mniejszym stężeniu do środowiska o stężeniu
większym, czyli wbrew gradientowi stężeń. Taki
transport wymaga dostarczenia energii. Jej źródłem
jest hydroliza cząsteczki ATP.
Przykładem mechanizmu transportującego jest
pompa sodowo-potasowa.
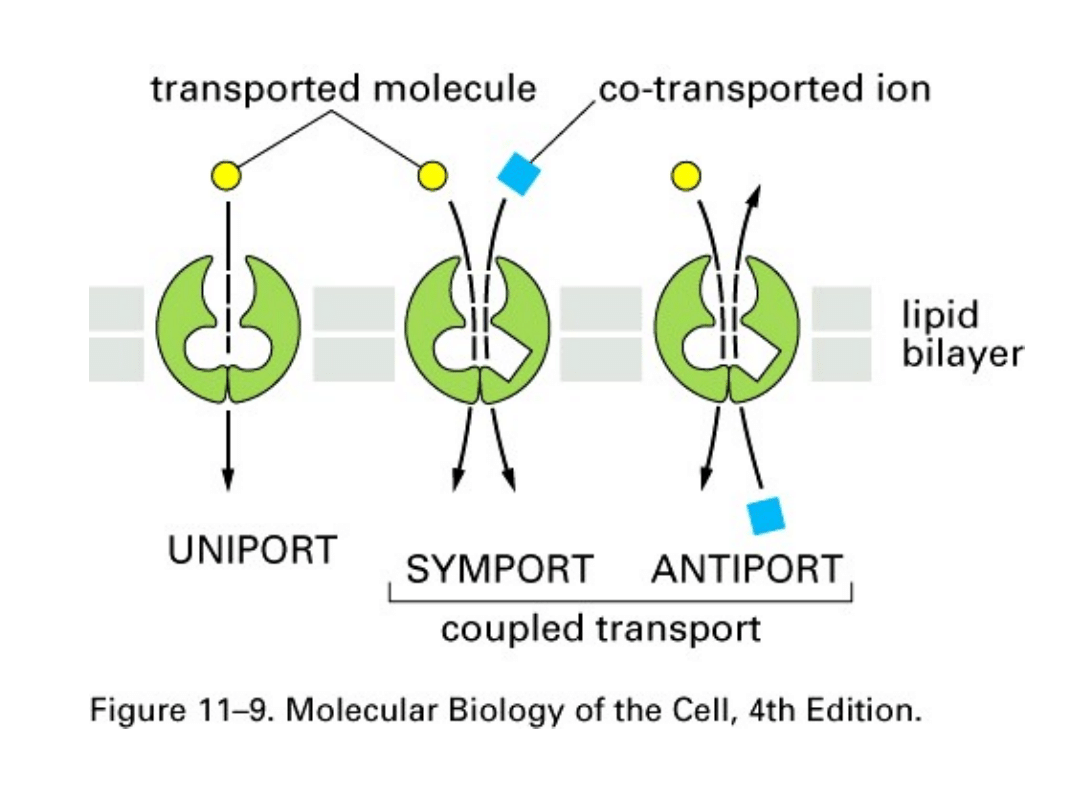
Aktywny transport substancji może być następujący:
• uniport
- transport jednej cząsteczki (substancji)
• symport
– transport dwóch substancji w tym samym
kierunku
• antyport
- transport dwóch substancji w różnym
kierunku.

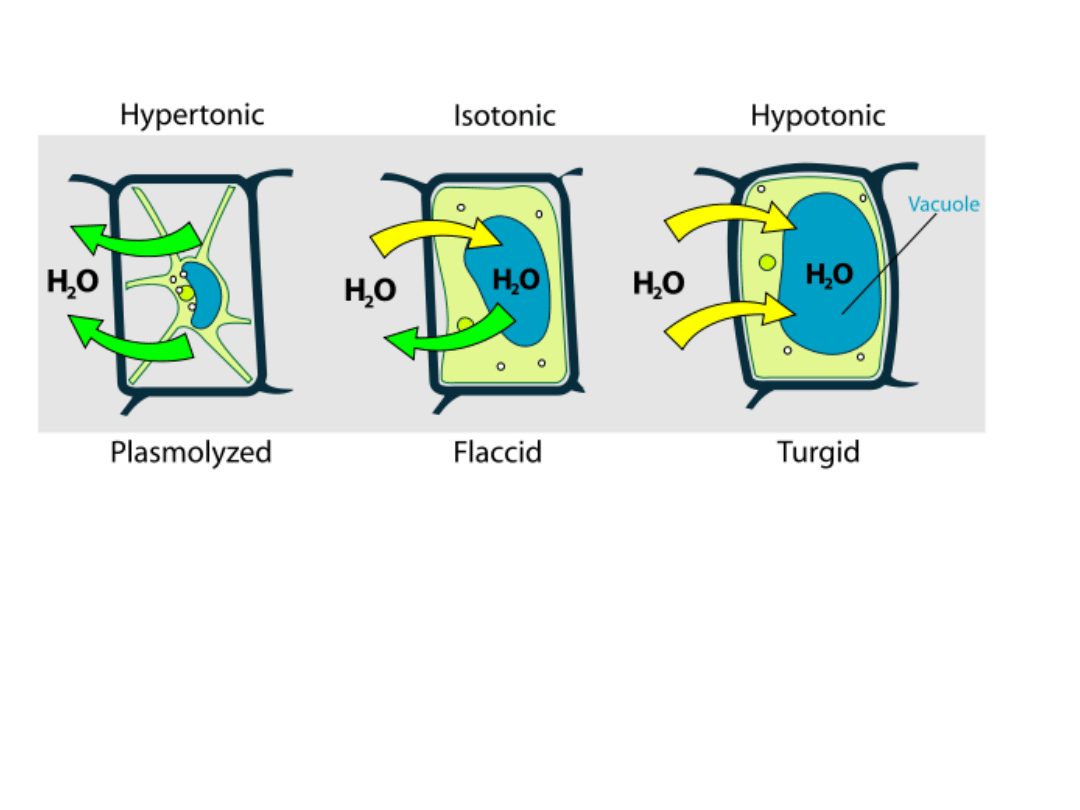
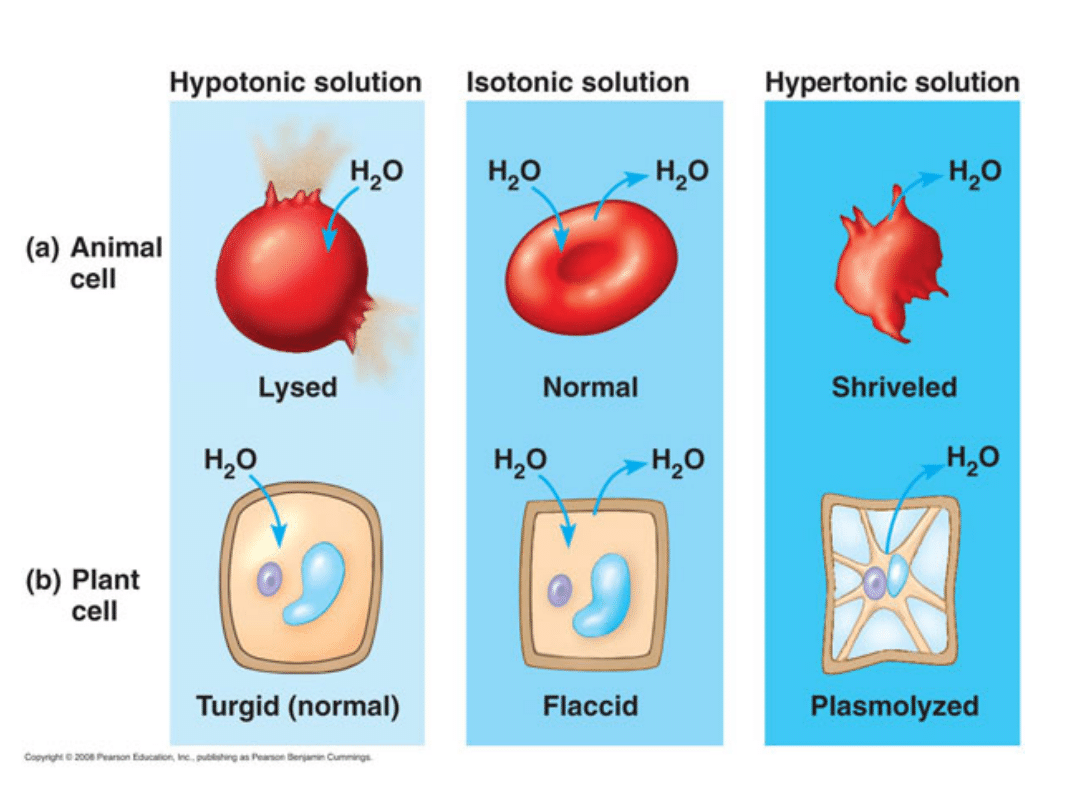
Roztwór izotoniczny - to roztwór, który posiada
identyczne ciśnienie osmotyczne w stosunku do
innego roztworu.
Aby roztwór wykazywał tę cechę, musi mieć
identyczne sumaryczne stężenie molowe wszystkich
związków zdolnych do przechodzenia przez błonę
półprzepuszczalną, przez które kontaktuje się z
innym roztworem. Przy kontakcie dwóch roztworów,
które są izotoniczne w stosunku do siebie, szybkość
osmozy w obu kierunkach jest identyczna, a zatem
układ osmotyczny pozostaje w równowadze
dynamicznej.
Pojęcie to jest najczęściej stosowane w biologii i
medycynie. Np. w medycynie płyny, które są
długotrwale podawane pacjentom przez kroplówkę,
muszą być izotoniczne, aby nie wywołać
odwodnienia organizmu.
Z tych samych przyczyn intensywnie trenujący
sportowcy piją płyny izotoniczne, gdyż większość
napojów tradycyjnych jest albo hipotoniczna albo
powoduje odwodnienie.

Roztwór hipertoniczny - roztwór, którego
ciśnienie osmotyczne jest wyższe od ciśnienia
osmotycznego innego roztworu, z którym go
porównujemy, przy czym zazwyczaj ciśnienie
osmotyczne omawianego roztworu odnosimy do
ciśnienia osmotycznego krwi lub innych płynów
ustrojowych człowieka. Zazwyczaj większe ciśnienie
osmotyczne ma roztwór posiadający większe
stężenie molowe, aczkolwiek bierzemy pod uwagę
jedynie stężenie cząstek zdolnych do przechodzenia
przez błonę selektywnie przepuszczalną (taką jest
np. błona komórkowa). Podanie człowiekowi (i w
ogóle organizmowi żywemu) roztworu
hipertonicznego powoduje plazmolizę komórek, co
może doprowadzić do ich śmierci.

Roztwór hipotoniczny - roztwór, który posiada
mniejsze ciśnienie osmotyczne od innego roztworu.
Aby roztwór posiadał tę cechę musi mieć mniejsze
stężenie związków chemicznych, które są zdolne do
przechodzenia przez błonę półprzepuszczalną,
przez którą kontaktuje się z innym roztworem.
Pojęcie to jest zazwyczaj stosowane w biologii i
medycynie. Komórki tkanki umieszczone w wodnym
roztworze hipotonicznym wchłaniają wodę i
nabierają objętości. W medycynie podawanie
kroplówki z płynu hipotonicznego jest stosowane w
szybkim zapobieganiu i leczeniu odwodnienia
organizmu.

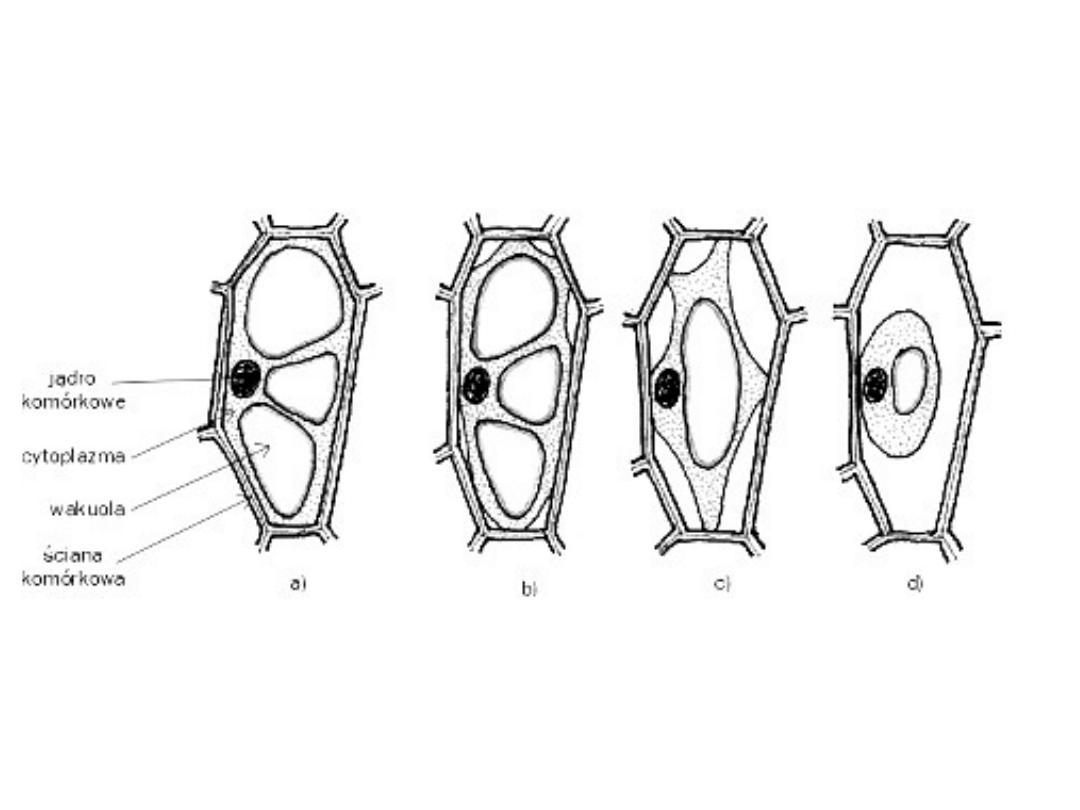
PLAZMOLIZA
Zjawisko odstawania protoplastu od ściany
komórkowej.
Plazmolizę można zaobserwować w komórkach
roślinnych po umieszczeniu ich w roztworze
hipertonicznym. Zgodnie z prawami osmozy woda
przenika przez plazmalemę z komórki do
roztworu, co powoduje odwodnienie komórki i
kurczenie się cytoplazmy podstawowej.
W zależności od stopnia utraty wody wyróżnia się
plazmolizę kątową, wklęsłą i wypukłą (graniczną).
Umieszczenie splazmolizowanej komórki w
roztworze hipotonicznym spowoduje napływ wody
do komórki i odzyskanie turgoru - deplazmolizę.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
- Slide 53
- Slide 54
Wyszukiwarka
Podobne podstrony:
Transport przez błony biologiczne, Studia, I rok, Wykłady z biofizyki
Błony biologiczne
Biochemia wykład 12 Błony biologiczne
wyklad 3 Transport przez blony biologiczne 1
54 Wlasciwosci blony biologicznej i blon molekularnych
2 b Transport lekow przez blony biologiczne
Błony biologiczne architektura i funckja sackman 1
12 BIOCHEMIA blony biologiczne
14. Transport cząsteczek przez błony biologiczne, Studia, biologia
Blony biologiczne id 75238 Nieznany (2)
błony biologiczne
Blony biologiczne K, dydaktyka, biologia-praktyki, II gimnazjum
BŁONY BIOLOGICZNE
BIOCHEMIA 5 Blony biologiczne
Błony biologiczne i transport błonowy
33 TRANSPORT BIERNY I TRANSPORT AKTYWNY JONOW SODU I POTASU PRZEZ BLONY BIOLOGICZNE
Błony Biologiczne
więcej podobnych podstron