
1
04.10.2007
METODY STOSOWANE W BIOLOGII KOMÓRKI
Biologia komórki:
Jest jedną z najszybciej rozwijających i zaawansowanych dziedzin przyrody
Współczesne badania koncentrują się na budowie, funkcji i zależnościami między
budową a funkcją
Komórki są podstawowym elementem każdego organizmu
Każda komórka otoczona jest błoną komórkową
Cechą charakterystyczna komórki jest zdolność do tworzenia własnych kopii
Ważnym wydarzeniem dla biologii komórki było wynalezienie mikroskopu
świetlnego w połowie XVII wieku; pozwoliło to odkryć komórkę
Przełomowe daty dla biologii komórki:
1655 – Robert Hook użył prymitywnego mikroskopu i jako pierwszy nazwał elementy korka
komórkami
1676 - Antonie van Leeuwenhoek przy użyciu mikroskopu odkrył pierwotniaki, a w 1685
odkrył bakterie
1833 - Brown obserwując storczyki opisał dokładnie jądro komórek roślinnych
1838 - Schwann i Schleiden ogłosili Teorię Komórkową - każdy organizm roślinny i
zwierzęcy składa się z komórek zawierających jądro komórkowe
1857 –
Kölliker opisał mitochondriom w komórkach mięśniowych
1873 - Golgi Camillo opracował metodę barwienia preparatów mikroskopowych solami
srebra, dzięki czemu zbadał budowę neuronu
1879 - Flemming opisał zachowanie się chromosomów mitotycznych w komórkach
zwierzęcych
1879 – Cayal opracował metody barwienia, które umożliwiły badania nad budową komórki
nerwowej
1898 - Golgi opisał poraz pierwszy struktury, które zostały nazwane od jego nazwiska
(aparaty Golgi'ego)
1902 - Theodor Boveri odkrył związek chromosomów z dziedzicznością
1952 - Pallad i inni rozwinęli metody mikroskopii elektronowej, pozwoliło to dostrzec
budowę cytoszkieletu i ultrastrukturę organelli
1957 - Robertson korzystając z mikroskopii elektronowej wykazał dwuwarstwowy charakter
błony komórkowej
STOSOWANE METODY BADAWCZE W BIOLOGII KOMÓRKI:
Badania mikroskopowe:
obserwacje mikroskopowe umożliwiają zapoznanie się z budową i funkcją oraz
analizą procesów zachodzących w komórkach
oglądać można komórki, organelle i cząsteczki
przeciętna średnica komórek wynosi od 5 do 20 mikrometrów
Mikroskop świetlny:
pozwala na powiększenie do 1000x i jest podstawowym narzędziem badawczym w
biologii komórki
zdolność rozdzielcza 0,2 mikrometrów

2
zdolności rozdzielcza
- najmniejsza odległość między dwoma dostrzegalnymi
punktami
aby oglądać komórki w mikroskopie świetlnym potrzeba 3 czynników:
odpowiednio przygotowany preparat (cienki i często wybarwiony)
padające światło skupione przy pomocy soczewek w kondensatorze
aby ogniskować obraz w oku potrzeba systemu dobrze ustawionych soczewek
Odmiany mikroskopu świetlnego:
kontrastowo-fazowy
kontrastuje struktury, które w normalnych mikroskopach są przejrzyste i słabo
widoczne
pozwala na przekształcenie przesunięć fali świetlnej,
zmiany jej amplitudy są lepiej widoczne przez oko ludzkie
(nie ma różnicy między nimi) - pozwala to na przesunięcie amplitudy
fali świetlnej, a obiekty stają się niewidoczne
interferencyjny
stwarza podobne możliwości – korzysta z interferencji (nakładania) - nakłada
wiązki świetlne na siebie
nie jest konieczne barwienie
układ optyczny morawskiego
– powszechnie stosowany do obserwacji nie
barwionych komórek;
fluorescencyjny
pozwala na uzyskanie informacji cytochemicznej, dzięki zastosowaniu UV
jako źródła światła, który wywołuje fluorescencję niektórych substancji
chemicznych
fluorescencja
o pierwotna/naturalna (chlorofil świeci na czerwono; kutyna na żółto-
zielono; lignina na żółto-pomarańczowo-zielono)
o
wtórna (występuje w przypadku stosowania
fluorochromów
-
specjalnych barwników fluoryzujących; oranż akrydyny łączy się z
DNA i powoduje świecenie się na zielono, RNA na czerwono, błękit
alininowy wykazuje żółtą fluorescencję kalozy)
konfokalny/współogniskowy
inne źródło światła - laser o zakresie fal bliskich UV
skanuje badany preparat, co pozwala wyeliminować warstwy znajdujące się
powyżej i poniżej podmiotu badanego
skanowany obraz jest zapisywany i wyświetlany
na ekranie monitora trójwymiarowej rekonstrukcji komórki
Mikroskopy elektronowe:
do tworzenie obrazu wykorzystuje się strumień elektronów o niewielkiej długości fali
umożliwia to milionkrotne powiększenie, przy rozdzielczości tysiąckrotnie mniejszej
niż w optycznym - 2nm
umożliwia wgląd do organelli, a także pojedynczych cząstek
ogólna budowa podobna do mikroskopu świetlnego:
działo elektronowe wytwarza strumień elektronów, przyspieszany w polu
elektromagnetycznym, naginany przez soczewki (wszystko w komorze
próżniowej)

3
W badaniach komórek wyróżniamy dwa typy mikroskopu elektronowego:
transmisyjny mikroskop elektronowy (TME)
o podobna zasada jak w mikroskopie świetlnym
o
przez preparat skontrastowany solami metali ciężkich przechodzi
strumień elektronów
o
obraz jest rzutowany na ekran, który jest pokryty warstwą substancji
fluoryzującej
o
pod wpływem bombardowania powstaje monochromatyczny obraz
utworzony przez jasne i ciemne kontury
o
rozdzielczość - 2nm;
scanningowy mikroskop elektronowy (SME)
o
tworzy wyraziste obrazy trójwymiarowe o dużej (3-20nm)
rozdzielczości;
o
strumień elektronów skanuje powierzchnie preparatu pokrytą przez
warstwę złota lub platyny
o elektrony są odbijane i wychwytywane przez detektor umieszczony po
wyżej preparatu, ten przekształca je w impulsy elektryczne
o
trudne i pracochłonne przygotowanie preparatu
o
po zobrazowaniu kształtu małych jednostek stosuje się barwienie
negatywowe, substancja wypełnia przestrzenie "pomiędzy"- powstają
jasne struktury na ciemnym tle
o
cieniowanie
- cienka warstwa platyny "pod kątem" pozwala na
pokazanie różnicy grubości napylanych struktur
o
metoda mrożenia i łamania
- badanie błon biologicznych. zamraża się
ciekłym azotem i cieniuje platyną .
Mikroskopia ta pozwala na zapoznanie się z morfologią lecz nie pozwala uzyskać
danych cytochemicznych
Badania cytochemiczne:
pozwalają na odkrycie in situ konkretnych związków i substancji chemicznych
reakcja PAZ (na wykrycie obojętnych wielocukrów)
reakcja Folgena (wykrywa kwasy nukleinowe)
reakcja FIF (fluorescencja indukowana formaldehydem, wykrywanie amin)
Metody immunocytochemiczne
rozpoznaje białka
wykorzystują wysokie swoistości wiązania się antygenu z przeciwciałem
cząsteczka przeciwciała wiąże i rozpoznaje konkretny antygen poprzez
podawanie konkretnych przeciwciał
Metoda hybrydocytochemiczna
wykrywanie DNA i RNA
wykorzystują zjawisko hybrydyzacji kwasów nukleinowych
rozpoznają konkretne sekwencje nukleotydów - odcinki komplementarne łączą
się in situ (w miejscu)
odpowiedni kwas musi mieć odpowiednią sekwencję
syntetyzuje się komplementarną strukturę - sondę, następnie znaczy się
fluorochromami, antygenami lub izotopami promieniotwórczymi
łączą się (hybrydyzują) i np. świecą - przy fluorochromie
FISH (fluorescent in situ hybridization) - pozwala na zlokalizowanie i
identyfikację

4
pozwalają na zidentyfikowanie obcych kwasów nukleinowych; jeśli jest mało
kwasu DNA, należy namnożyć DNA oraz zastosować PCR in situ (techniką
reakcji łańcuchowej polimerazy) - następuje powielenie badanego odcinka
Metoda autoradiografii
pozwala na wykrycie izotopów promieniotwórczych
wykorzystując ich zdolność wybarwienie się emulsji światłoczułych
stosowanych w fotografii (na czarno)
po wprowadzeniu substancji zawierającej izotopy, można śledzić procesy
metaboliczne(znakowanie substancji)
pokrycie gotowego preparatu cienką warstwą substancji światłoczułej
promieniowanie beta powoduje wytrącenie się z azotanu srebra - ziarna
srebra, które uwidocznione są na preparacie
na preparacie w miejscu znaczenia występują stronty w miejscach określonych
syntez oraz można określić ich aktywność przez ilość strontów
służy do oznaczenia białek i kwasów nukleinowych
Cytometria przepływowa
metoda ta pozwala na ilościową ocenę właściwości fizycznych i
chemicznych pojedynczych komórek przepływających przez miejsce pomiaru
(tylko komórki w formie zawiesiny - m.in. w formie wyizolowanych jąder
<ploidalność, DNA>
odczyty z każdego pomiaru są zliczane, rejestrowane i statystycznie
analizowane przez komputer
nie należy do technik mikroskopowych, jednak przygotowanie obejmuje
metody cytochemiczne i izometryczne
bardzo szerokie zastosowanie w medycynie i weterynarii
można badać kształt, rozmiar i zawartość DNA, ploidalność, obecność gazów,
białek, cukrowców, przepuszczalność błon, aktywność błon, wewnętrzne pH
komórki oraz stężenie jonów
w praktyce klinicznej służy do diagnostyki chorób krwi, zaawansowania
chorób nowotworowych, zaburzeń układu immunologicznego
Badania biochemiczne i molekularne:
mają na celu wyizolowanie z komórek, a następnie scharakteryzowanie określonych
substancji chemicznych w szczególności białek i kwasów nukleinowych (ex situ -
poza komórką)
do rozdziału białek i kwasów nukleinowych służy
elektroforeza
metoda ta wykorzystuje zdolność do poruszania się cząstek obdarzonych ładunkiem w
polu elektrycznym
służy do rozdziału mieszanin białek i kwasów nukleinowych, które różnią się
ładunkami, więc i prędkością poruszania się jonów w polu elektrycznym
po pewnym czasie elektroforezy następuje rozdział mieszanin wzdłuż linii pola
elektrycznego
każdy ze składników znajduje się w innym miejscu podłoża stałego
w ten sposób, wycinając odpowiedni fragment podłoża stałego można wyizolować
skład
Podłoża:

5
do kwasów nukleinowych – elektroforeza na żelach agarowych (kwasy
nukleinowe wcześniej pocięte na krótsze odcinki przy użyciu enzymów
restrykcyjnych, działających w odpowiednich miejscach)
do białek – elektroforeza na żelu poliakrylamidowym ( w obecności SDS -
dodecylosiarczanu sodu)
Największą efektywność w rozdzielaniu białek wykazuje
elektroforeza
dwukierunkowa
(rozdział na dwukierunkowej płaszczyźnie). Pozwala na
wyodrębnienie 1000 białek z jednej próbki
Do identyfikacji (sekwencjonowania) rozdzielonych białek i sekwencjonowania
kwasów nukleinowych najczęściej służy
blotting
(odciskanie, plamy na bibule):
suthern blotting (południowy) – wykrywa pojedyncze, krótkie odcinki DNA
i RNA, stosowana jest sonda (komplementarna do odcinka kwasu
nukleinowego – DNA lub RNA)
northern blotting (północny) - tylko RNA, też przy udziale sond
western blotting (zachodni) - wykrywa białka przy użyciu odpowiednio
znakowanych przeciwciał
Rozdzielane podczas elektroforezy substancje przenosi się z żelu na błonę
nitrocelulozową lub nylonową (polega na odciśnięciu w obecności roztworu
ekstrakcyjnego, z żelu na odpowiednią błonę)
Zachowuje rozmieszczenie rozdzielonych elementów i sekwencjonowanie przy użyciu
metody hybrydyzacji przy pomocy sond lub przeciwciał znakowanych enzymami,
pierwiastkami promieniotwórczymi (izotopami)
Są bardzo czułe - pozwalają na wykrycie już od 10 pikogramów substancji
PCR (Polymerase Chain Reaction)
- umożliwia wykrycie śladowych ilości kwasów
nukleinowych, polega na powieleniu cząsteczek kwasów nukleinowych
11.10.2007
BUDOWA I RÓŻNICOWANIE SIĘ KOMÓREK
komórka roślinna i zwierzęca 5-20 mikrometrów
wszystkie komórki powstały w toku ewolucji zjednej prakomórki wyjściowej (ok. 3
mld lat temu);
Podział:
Prokaryota:
komórki bez jądra komórkowego
bez organelli
bakterie i sinice (cyjanobakterie)
bardzo małe < 7 mikrometrów
w większości jednokomórkowe, ale zorganizowane w grona, łańcuchy etc
nie posiadają błony, wyodrębnionego jądra ani organelli
występuje gruba ściana komórkowa (15-100 mikrometrów)
ściana komórkowa zbudowana z proteoglikanów; czasem pokryta otoczką
śluzową
u bakterii chorobotwórczych ściana komórkowa zbudowana z cząsteczek
glikoproteinowych
ściana komórkowa utrzymuje kształt i chroni przed rozerwaniem
właściwa błona pod ścianą jest kompleksem białkowo-lipidowym

6
tworzy liczne wypustki do wnętrza komórki, prawdopodobnie zwiększając
powierzchnię oddechową bakterii
zawiera również liczne receptory białkowe - w ten sposób może dostosować
się do warunków środowiska
chromosom jest kolistą dwuniciową cząsteczką DNA
w cytoplazmie znajdują się mRNA, tRNA i rRNA
rybosomy ze stałą sedymentacji 70s (biosynteza białka)
liczne pęcherzyki gazowe, krople lipidów, wielocukry, nieorganiczne
fosforany, zorganizowane w postaci ziaren sferycznych
wiele bakterii posiada zdolność ruchu, zbudowane przez rzęski z flagelliny
elementy roślinneto gruba ściana komórkowa
elementy zwierzęce to rzęski zbudowane z flagelliny
cyjanobakterie (sinice)
o
najprymitywniejsze autotrofy zawierające chlorofil
o
zamieszkują wszystkie nisze ekologiczne
o barwniki : chlorofil, karoten, ksantofil, fitocyjanina i tzw. skrobia
sinicowa
o
są one rozpuszczone w cytoplazmie lub wbudowane w błoniaste
struktury (nie posiadają chloroplastów)
o
bakterie i cyjanobakterie zawierają złożone układy, które umożliwiają
im syntezę wszystkich niezbędnych składników
o
potrafią wiązać N i C
o
stanowią główną masę biosfery
Eukaryota:
posiadają komórki eukariotyczne z wyodrębnionym jądrem komórkowym oraz
organellami
występują zarówno jednokomórkowe (drożdże i pierwotniaki) jak i
wielokomórkowe (grzyby, rośliny i zwierzęta)
posiadają: mitochondria, chloroplasty, cytoszkielet, lizosomy, rybosomy,
aparat Golgiego
JĄDRO:
o
największa struktura, położona centralnie
o stanowi centrum informacji genetycznej
o chromatyna w chromosomach
MITOCHONDRIA:
o
podwójna błona
o owalne
o
posiadają własne DNA
o
potrafią się dzielić
o magazynowana energia w postaci wysokoenergetycznych wiazań ATP
CHLOROPLASTY:
o
występują u roślin
o
podwójna błona
o
własne DNA
o chlorofil potrzebny do fotosyntezy
BŁONY:
o
siateczka śródplazmatyczna - gładka lub szorstka
LIZOSOMY :
o System GERLL

7
o
Enzymy proteolityczne, tworzące wewnątrzkomórkowe
PEROKSYSOMY:
o
Środowisko do rozkładu nadtlenku wodoru
CYTOZOL:
o
wewnątrzkomórkowa przestrzeń
o
wodny żel, w którym przebiegają reakcje chemiczne (synteza białek,
rozkład cząsteczek pokarmowych)
CYTOSZKIELET:
o
system włókien - system włókien: filamenty i mikrotubule, które
rozciągają się przez całą komórkę
o
decyduje o wewnętrznej organizacji komórki
o
niezbędny w komórkach zwierzęcych i roślinnych
o system rurek i beleczek nadaje odporność mechaniczną
o
umożliwia ruch
o
warunkuje kształt
o bierze udział w podziałach komórkowych – organizacja wrzeciona
kariotycznego i cytokinetycznego
Jedność i różnorodność komórek:
Różnice:
wszystkie komórki różnią się rozmiarem i kształtem
bakteria bdellowibria ma kształt torpedy, która porusza się do przodu przez owiniętą
wić
komórki:
zwierzęce - błona komórkowa
roślinne - ściana komórkowa
tkanki kostne - otoczone twardą zmineralizowaną substancją
również funkcje różnią komórki
tkanka każdego narządu pełni inne funkcje
komórki różnią się aktywnością
niektóre pobierają tlen, dla innych jest on letalny
produkują hormony, enzymy, skrobie, tłuszcz i barwniki
kształt, wielkość, funkcje i wymagania różnią komórki
wszystkie komórki podobne pod względem biochemicznym, mają podobną
ultrastrukturę
Podobieństwa:
wszystkie składają się z genów zapisanych trójkowym kodem chemicznym
replikują - synteza białek taka sama w każdej komórce
białka składają się z takich samych (23) aminokwasów, które połączone są w różnych
sekwencjach
Prakomórka powstała 3-3,8 mld lat temu - ulegała ewolucji na skutek mutacji
ORGANIZMY MODELOWE (RÓŻNICOWANIE OD KOMÓREK
PROKARIOTYCZNYCH DO EUKARIOTYCZNYCH)
BAKTERIE:
najmniejsze i najprostsze prokatyota
zróżnicowany kształt (kulisty, pałeczkowaty, śrubowaty)
długość kilka mikrometrów
gruba ściana komórkowa
błona komórkowa

8
w centrum pojedyncza lub podwójna cząsteczka DNA
w mikroskopie elektronowym wnętrze nie zawiera zorganizowanej struktury i
organelli komórkowych
rozmnażają się przez podział z częstotliwością co 20 min, także jedna bakteria może w
ciągu 11 h wytworzyć 15 mld komórek
stąd też mogą szybko ewoluować, zdobywać nowe źródła pokarmu
zdobywają odporność na antybiotyki
wykorzystują ogromną rozpiętość środowiska - od wulkanów do wnętrza innych
komórek
niektóre fotosyntetyzują, inne chemosyntetyzują
pod względem ilościowym przewyższają inne organizmy
modelowa bakteria - Escherichia coli – żyje w jelicie cienkim, posiada 4000 białek z
DNA; pierwsze eksperymenty na kwasach nukleinowych; replikacja i odczytywanie
przebiega jak u wyżej zorganizowanych organizmów
GIARDIA:
prymitywny eukaryota
pośredni etap w ewolucji
jednokomórkowy organizm pasożytniczy
posiada dwa wyodrębnione identyczne jądra
nie posiada organelli ani struktur - poza cytoszkieletem
pasożytuje w warunkach beztlenowych w jelicie cienkim
DROŻDZE PIEKARSKIE:
typowe komórki eukariotyczne
małe jednokomórkowe grzyby
otoczone grubą ścianą komórkową (cecha charakterystyczna dla świata roślin)
zawierają mitochondriom
nie posiadają chloroplastów
rozmnażanie przez podział tak szybko jak bakterie
PIERWOTNIAKI:
jednokomórkowe, ale w wielu formach duże i złożone
bardzo agresywne
ruchliwe bądź osiadłe
mięsożerne lub fotosyntetyzujące
posiadają wyrostki czuciowe, fotoreceptory, rzęski, aparaty gębowe, wiązki kurczliwe
podobne do mięśni
DIDINUM:
bardzo podobna budowa do organizmów wielokierunkowych
duży
mięsożerny
do 150 mikrometrów
kulisty kształt
otoczony dwoma pasmami rzęsek
uwypuklenie podobne do ryjka
szybko pływa i odżywia się innymi pierwotniakami wyrzucając z ryjka paraliżujące
strzałki

9
OEGANIZMY WIELOKIERUNKOWE:
grzyby
rośliny
zwierzęta
Arabidopsis thaliana – rzodkiewnik pospolity – modelowa roślina spośród 300000
roślin
niewiele DNA 3-4 razy więcej niż u drożdży
z tej uwagi jest rośliną modelową w badaniach genetycznych i biochemicznyc
Zwierzęta:
muszka owocowa, nicień, mysz
spośród wielokomórkowych, większość stanowią owady (Drosophila
melanogaster)
modelowy nicień (Caenorhabditis elegans)
o jest mniejszy i prostszy niż Drosophila melanogaster
o
dokładnie poznano już rozwój od zapłodnionej komórki jajowej do
dojrzałej postaci
o
składa się z 959 komórek u dojrzałego osobnika
Homo sapiens
o badane jedynie w hodowlach komórek z powodów etycznych
o
dokładnie poznane mutacje w tysiącach genów
o
znana kolejność nukleotydów i liczbą genów, ale nie znane dokładne
funkcjonowanie
o
duże zróżnicowanie komórek w tkanki, te z kolei zorganizowane w
narządy
o
komórki różnych tkanek są bardzo niepodobne (np. tłuszczowa i
kostna)
o
powstają w rozwoju embrionalnym z jednej zapłodnionej komórki
jajowej
o
wszystkie mają jednakowe DNA
o
różnicują się poprzez różne wykorzystywanie sekwencji
o
nie wszystkie geny są wykorzystywane we wszystkich komórkach
o
różna ekspresja genów w różnych komórkach
o
posiadają identyczne instrukcje genetyczne, ale w różny sposób
wykorzystywane, dlatego są tak zróżnicowane (nie całe DNA jest
wykorzystywane)
18.10.2007
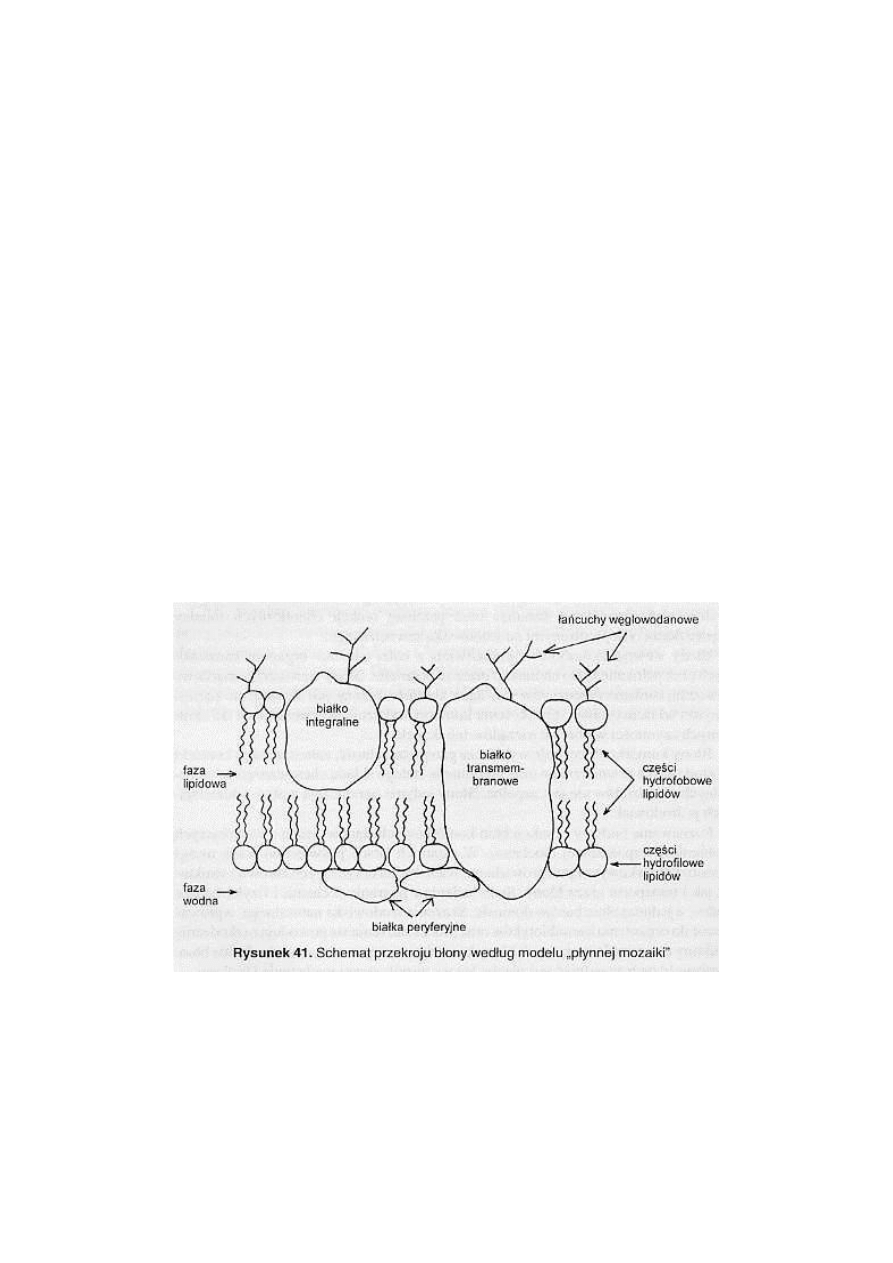
BŁONY KOMÓRKOWE
Podstawowy składnik każdej komórki
Stanowią barierę oddzielającą zawartość komórki od środowiska i od siebie
Umożliwiają wzrost i zmianę kształtu
Selektywne bariery między przestrzeniami
Bakterię mają pojedynczą błonę (oprócz błony, otoczone są cytolemmą, ale zawierają
stosunkowo dużo błon wewnętrznych – 95% - 98% wszystkich błon)
Wyróżniamy:
Błonę jądrową (nukleolemmę)
10
Błonę siateczki śródplazmatycznej (retikulum)
Błonę układu Golgi’ego
Błony lizosomalne
Błony mitochondrialne
Błony innych wewnątrzkomórkowych elementów obłonionych
Budowa:
Wszystkie błony zbudowane są podobnie
Białka błonowe nadają charakter błonom
Skład:
Składają się z dwóch podstawowych elementów: białek i lipidów
Niektóre posiadają jeszcze elementy węglowodanowe
Wspólny plan budowy – modele:
Płynna mozaikowa budowa błon, zakłada że składają się z podwójnej warstwy
lipidowej, w której zanurzone są cząsteczki białka
Podwójna błona, zbudowana z
hydrofilowych
cząsteczek znajdujących się na
zewnątrz oraz z nierozpuszczalnych
hydrofobowych
cząsteczek znajdujących się
wewnątrz
Lipidy błonowe są
amfipatyczne
– mają hydrofilowe i hydrofobowe właściwości
Posiada zdolność do tworzenia dwucząsteczkowej warstwy, w odpowiednich
warunkach
liposomów
– kulistych twórów
Właściwości:
Względna ciągłość (posiada tylko warstwa lipidowa) – cząsteczki warstwy lipidowej
mogą się rozsuwać i obejmują agregaty lub cząsteczki białkowe (białka błon nie
stanowią fazy ciągłej)
Białka błon posiadają
labilność
, co umożliwia zmianę ich lokalizacji i występowanie
w różnych miejscach błon, co z kolei prowadzi do lokalnej specjalizacji błony

11
Lipidy i białka błon posiadają zdolność zmiany położenia wzdłuż płaszczyzn błon (z
jednej warstwy do drugiej –
ruchy flip-flop
) –
dyfuzja błon
Mozaikowość
– rozmieszczone białka i lipidy w podwójnej warstwie lipidowej jest
asymetryczne – brak powtarzających się jednostek (jedna strona błony różni się
składem chemicznym od drugiej)
Właściwości lipidów błonowych:
1. Fosfolipidy
Aminowe – fosfatydyloetanoloamina, fosfatydyloseryna
Cholinowe – fosfatydylocholina, fosfatydyloinozytol, fosfatydylosfingozyna
2. Glikolipidy
Glikosfingolipidy
Gangliozydy
galaktocerebriozy
3. Cholesterol
Lipidy błonowe:
Tworzą w wodzie dwuwarstwy – uniwersalna podstawa budowy błon komórkowych
Hydrofilowa główka z rozpuszczalną w wodzie grupą polarną
Hydrofobowe ogonki z grupami nie polarnymi, tworzą wiązania z tłuszczami
W wyniku tych sprzecznie działających sił tworzy się dwuwarstwa
Dwuwarstwa lipidowa ma zdolność samozasklepiania się - w przypadku rozdarcia,
spontanicznie przegrupowuje się dwuwarstrwa w celu zasklepienia
Małe rozdarcie – reorganizacja prowadzi do naprawy
Duże rozdarcie – warstwa rozpada się na samodzielne pęcherzyki
Dwuwarstwa lipidowa jest dwuwymiarowym płynem
Wodne środowisko uniemożliwia ucieczkę lipidów błonowych z dwuwarstwy, ale
mogą się przemieszczać (dlatego zachowuje się jak dwuwarstwowy płyn)
Liposomy – (maja średnicę około 25 nanometrów do 1 milimetra)
W normalnych warunkach dwuwarstwa jako dwuwymiarowy płyn, w którym
cząsteczki lipidowe maja swobodny ruch (obrotowe ruchy), mogą przeskakiwac z
jednej warstwy do drugiej (ruchy flip-flop)
Płynność błony:
Umożliwia szybką dyfuzję białek błonowych w płaszczyźnie dwuwarstwy i wzajemne
oddziaływanie ważne w sygnalizacji komórkowej
Umożliwia fuzję błon ze sobą i wymieszanie ich cząsteczek, ważne przy podziałach –
równomierne rozdzielenie pomiędzy komórkami potomnymi
W komórkach zwierzęcych płynność jest zmieniana dzięki obecności steroidu –
cholesterolu
Cholesterolu nie posiadają drożdże, rośliny i bakterie
Cholesterol – krótkie, sztywne cząsteczki w dużych ilościach, w przestrzeniach
pomiędzy sąsiednimi cząsteczkami fosfolipidów usztywniają dwuwarstwę i
zmniejszają ich przepuszczalność
Dwuwarstwa lipidowa jest asymetryczna, a wynika to z:
Nierównomiernego położenia fosfolipidów po obu stronach
Nierównomiernego ułożenia glikolipidów (występują tylko po zewnętrznej
stronie)
Przewagi pewnej zawartości białek integralnych i powierzchniowych po
stronie wewnętrznej
Selektywne przepuszczanie (dyfuzja)

12
Błony plazmatyczne przepuszczają pewien rodzaj cząsteczek, a inne
zatrzymują
Stanowią barierę ochronną
Główna rolę odgrywa hydrofobowe wnętrze – bariera zapobiegająca przejściu
większości cząsteczek hydrofilowych
Niektóre substancje przenikają przez
dyfuzję
(najszybciej, gdy są najmniejsze
i im łatwiej rozpuszczają się w olejach; są to małe i nie polarne cząsteczki –
tlen, dwutlenek węgla, benzen; szybko również przechodzą nierównomierne
cząsteczki – woda, etanol; wolniej przechodzi glicerol)
Błony są nie przepuszczalne dla jonów i substancji nierozpuszczalnych w
wodzie
Większe cząsteczki nie przechodzą na zasadzie dyfuzji – glukoza,
aminokwasy, nukleotydy
Białka błonowe:
Większość funkcji jest pełniona przez konkretne białko
Różnią się ciężarem cząsteczkowym, budową (od 1000 do kilku tysięcy daltonów)
Związki białka stanowią 50% masy większości wszystkich błon
Ze względu na lokalizacje wyróżniamy:
Białka strukturalne-integralne, związane z błoną
Białka powierzchniowe (peryferyczne) nie są bezpośrednio związane z błoną
Ze względu na funkcję wyróżniamy:
Białka transportujące – pompa sodowo-potasowa (sód na zewnątrz, a potas do
wewnątrz
Białka wiążące – integryny wiążą wewnątrzkomórkowe filamenty aktyny z
białkiem substancji zewnątrzkomórkowych
Receptory – receptor płytkopochodnego czynnika wzrostu, wiąże
zewnątrzkomórkowe PGDF i wytwarza wewnątrzkomórkowe sygnały
inicjujące wzrost komórki
Enzymy – cykloza adenylanowa
Białka błonowe:
Mogą być związane z dwuwarstwą lipidową:
Bezpośrednio – są to transbłonowe białka integralne
Peryferycznie – poza obrębem dwuwarstwy, są tylko zakotwiczone grupami
lipidowymi
Pośrednio (powierzchniowe) – po jednej lub po drugiej stronie związane za
pośrednictwem innego białka
Łańcuch polipeptydowy zazwyczaj przechodzi przez dwuwarstwę jako alfa-helisa
(przechodzi tylko raz)
Wiele białek błonowych jest receptorami sygnałów zewnątrzkomórkowych
Białka tworzą
pory wodne
Jak można usunąć białka:
Integralne związane z błoną mogą być uwolnione poprzez detergenty
Luźno związane z błona można uwolnić w procesie ekstrakcji nie uszkadzając
dwuwarstwy lipidowej
Roślinne białka błonowe:
Są mało poznane

13
W szpinaku znajduje się około 100 odmiennych polipeptydów (80% to integralne, a
20% to powierzchniowe)
Związane są z cytoplazmatyczna warstwą
Najlepiej poznane białka błonowe:
Pompa protonowa i wapniowa
Nośnik jonów fosforowych i azotanowych
Nośniki sacharozy i auksyny
Kanały potasowe i wodne (akwaporyny)
Receptory etylenu i cytokininy
Grupa kinaz białkowych
Związki białek błonowych (modelowy przykład – erytrocyty – białka erytrocytów):
1. Glikoforyna A
Transblonowe
Monotypowe
Tworzą pojedyncze bloki
Masa cząsteczkowa - 28 kilodaltonów
Składają się z 131 reszt aminokwasowych silnie zglikolizowanych
60% łańcucha związana jest ze 100 resztami cukrowymi (wystają ponad
warstwę lipidową
Do głównych reszt cukrowych glikoforyny należy kwas sialowy
W jednym erytrocycie glikoforyna występuje 6 * 10
5
kopii
Erytrocyty bez tego białka nie wykazują zaburzeń fizjologicznych
2. Trzeciego szczytu elektroforetycznego B3SE
B3SE występuje około 6 * 10
6
kopii
Politopowe
– tworzy wielokrotne bloki
Masa cząsteczkowa wynosi 100 kilodaltonów
Zbudowane z 929 reszt aminokwasowych
Wielokrotnie przechodzi przez ciało grubości błony nawet do 10 razy
Transbłonowe- reguluje transport aminokwasów podczas usuwania dwutlenku
węgla z erytrocytów
Białka dobrze poznane u bakterii:
Bakteriorodopsyna – małe białko, zbudowane z 250 aminokwasów, występujące w
dużych ilościach w błonach bakterii słonolubnych
Siedem razy przechodzi przez ciało długości politopowe
Wchodzi w skład receptorów
Retinol – nadaje ciemną fioletową barwę
Białka szkieletowe:
Białka błonowe tworzące pod błoną włóknistą wyściółkę
Kotwiczą się do niektórych transbłonowych
Utrzymują kształt komórki
Zapewniają elastyczność i wytrzymałość
Wyróżniamy 11 rodzajów białek szkieletowych
Spektryna
Składa się z dwóch monomerów alfa- i beta- helikalnie splecionych
Masa cząsteczkowa alfa- wynosi 240 kilodaltonów, natomiast beta- 22o
kilodaltonów

14
Oba dimery wiążą się ze sobą tworząc filamenty
Długość 100 nanometrów
Końce pozostałych tetramerów łączą się z filamentami cytoszkieletu (aktyna i
propomiozyna)
Nadaje kształt erytrocytów jako dwuwklęsłych dysków
Zaburzenia w ekspresji alfa- i beta-spektryny prowadzi do zaburzeń metabolicznych –
do zmiany kształtu i zaburzeń w transportowaniu
Maja podłoże genetyczne
Funkcje białek błonowych:
Przyczepiają filamenty cytoszkieletu do błony komórkowej
Przyczepiają komórki do macierzy międzykomórkowej
Przenoszą cząsteczki do komórki i z komórki
Działają jako receptory dla sygnalizacji
Wykazują swoistą aktywność enzymatyczną
Cukrowe składniki błon komórkowych:
Węglowodany stanowią od 2% - 10% masy błon
Występują jako cukrowce, związane końcami z białkami błon –
glikoproteiny
, z
końcami lipidów –
glikolipidy
Długie łańcuchy polisacharydów –
proteoglikany
Występują po zewnętrznej stronie błony
Poznano około 100 monosacharydów
Galaktoza
Mamoza
Glukoza
Galaktozamina
Glukozamina
Kwas sialowy
W polipeptydach kwasy połączone są luźno
Cukry mogą się łączyć różnymi typami wiązań z różnymi aminokwasami tworząc
rozgałęzienia
Tworzą
glikokaliks
związany na powierzchni komórek (zwłaszcza nabłonkowych)
Glikokaliks
występuje tylko w komórkach zwierzęcych
Glikokaliks:
Zabezpiecza przed uszkodzeniem mechanicznym
Silnie pochłania wodę (powierzchnia jest ślizka)
Wiele komórek (erytrocyty) mogą się prześlizgiwać przez szczeliny ścian naczyń
włosowatych
Zabezpiecza (zapobiega) zlepianiu się erytrocytów i przyczepianiu do ścian naczyć
krwionośnych
Powoduje asymetrie błony, ponieważ występuje tylko po zewnętrznej stronie błony
U roślin znajduje się
celuloza
– zasadniczy element nadający kształt komórkom roślinnym
Węglowodany białkowe:
Występowanie monosacharydów w końcach glikosacharydowych przyczynia się do
określenia grupy krwi
Kwas sialowy odpowiedzialny jest za ujemny ładunek elektryczny

15
Oligosacharydy pomagają w utrzymaniu pozycji glikoprotein i 4 rzędowe białka
globularnego
Rozpoznanie i przyleganie do innych komórek
Powstawanie odczynów zapalnych we wczesnych stadiach infekcji bakteryjnej
Rozpoznawanie wzajemne komórek podczas kolejnych etapów rozwoju organizmu
25.10.2007
TRANSPORT PRZEZ BŁONY
Funkcjonowanie komórek jest uzależnione od wymiany materii
Barierą ograniczającą jest otaczająca błona komórkowa
Dwukierunkowy transport
– błona komórkowa musi pobierać substancję potrzebne
do rozwoju jak i również usuwać produkty przemiany materii
Typy transportu:
Transport pojedynczych cząsteczek:
Bierny
Wynika z różnicy gradientów stężeń po obu stronach błony i odbywa się
zgodnie z gradientem – od wyższego do niższego
Szybkość transportu nie naładowanych cząsteczek nie zależy od różnicy stężeń
cząsteczek obdarzonych ładunkiem, zależy od różnicy stężeń i od gradientu
elektrochemicznego
Przez błonę ujemnie naładowaną szybciej będą transportowane cząsteczki
naładowane dodatnio, a ujemnie najmniej
Dyfuzja prosta
o Ze względu na hydrofobowy półprzepuszczalny charakter błony,
swobodnie przenikają do wnętrza w drodze dyfuzji prostej, związki
dobrze rozpuszczalne w lipidach oraz dwutlenek węgla, tlen i azot
(cząsteczki gazowe), mocznik, etanol, glicerol, woda
o
Transport ten odbywa się bez udziału przenośników
o Natomiast dyfuzja ułatwiona uzależniona jest od przenośników
(Białka transbłonowe, które stanowią przejśccia do poszczególnych
substancji)
o
Przez błonę przechodzą jony, drobnocząsteczkowe związki organiczne
takie jak glukoza
o
Zestaw białek transportujący w błonie komórki lub organelli
wewnątrzkomórkowych ściśle określa co może wejść lub wyjść
Każdy typ błony ma swój własny zestaw białek transportujących:
o
Białka nośnikowe
Przejściowo łączą jony lub cząsteczki transportowe w ten
sposób, ze zmieniają swoja konformację przenosząc je na
druga stronę błony
W ten sposób transportowane są małe cząsteczki organiczne i
nieorganiczne, jony
Każdy przenośnik transportuje jeden typ cząsteczek
o
Białka kanałowe
są najczęściej zamknięte
mogą otwierać się pod wpływem trzech elementów
1. zmiany ąłdunku elektrycznego błony

16
2. związanie z ligandem (informator I lub II rzędu)
3. pod wpływem czynników mechanicznych
tworzą hydrofilne pory przez które przechodzą substancje na
drodze dyfuzji
większość kanałów przepuszcza tylko jony nieorganiczne,
dlatego nazywane są
kanałami jonowymi
Aktywny
Cząsteczki przechodzą przez błonę przeciwko gradientowi stężeń
Transport substancji odbywa się od mniejszego do większego stężenia
Czynny transport, energochłonny, związany z określonymi procesami
chemicznymi, w których biorą udział białka wchodzące w budowę ATP-azy
Transportowane są jony sodu, potasu i wapnia, poprzez pompę sodowo-
potasową i pompę wapniową
Pompa sodowo-potasowa:
Polega na transporcie jonów sodu na zewnątrz komórki, a jonów potasu do wnętrza
komórki
Ma na celu utrzymanie różnic stężeń obu jonów występujących pomiędzy wnętrzem a
przestrzenią międzykomórkową
Proces zachodzi przy udziale energii z hydrolizy ATP-azy
Cząsteczka transbłonowej ATP-azy jest związana na wewnętrznej powierzchni błony
Hydrolizuje ATP do ADP i ortofosforanów
Ulega fosforylacji
Jednocześnie wiąże się na swojej powierzchni jony sodu
Fosforylacja zmienia konformację ATP-azy powodując jej otwarcie na zewnątrz
powierzchni błon
Jon sodu jest uwalniany
Z ATP wiąże się jon potasu
Jednocześnie następuje defosforylacja
Zmienia się konformacja
W wyniku każdej zmiany na zewnątrz transportowane są trzy jony sodu, a do wnętrza
dwa jony potasu
Występuje w błonie komórek nerwowych i jest podstawą depolaryzacji i
hiperpolaryzacji, a zatem i jej pobudzania i hamowania
Działa w błonie komórek nabłonkowych, jelita, nerek, gruczołów śluzowych,
potowych, splotu naczyniowego
Wiąże się z mechanizmem wydzielania i usuwania moczu, śliny, potu, płynu
mózgowo-rdzeniowego
Kontroluje objętość komórki, bo podążająca za jonami sodu i chloru woda powoduje
hipotonię i pęcznienie komórki (zapobiega temu pompa sodowo-potasowa)
Pompa wapniowa:
Transport wbrew gradientowi stężeń przez transbłonowe białko
Umożliwia skurcz włókna mięśniowego
Energię potrzebną do transportu czerpie z hydrolizy ATP wewnątrz włókna
mięśniowego
Odbywa się tu transport oscylacyjny w dwóch kierunkach
Z siateczki śródplazmatycznej do sarkoplazmy –
skurcz
Z sarkoplazmy do siateczki śródplazmatycznej –
rozkurcz
Odpowiednie stężenie jonów wpływa na podziały komórkowe (mitoza)

17
Apoptoza (śmierć komórkowa zaprogramowana)
Wtórny przekaźnik informacji
Transport makrocząsteczek:
Większe cząsteczki oraz bakterie, fragmenty komórek nie mogą być transportowane
na zasadzie wyżej wymienionych
Ale mogą być wprowadzone poprzez wpuklenie –
endocytoza
Endocytoza:
Transport makrocząsteczek w postaci obłonionych cząsteczek z zewnątrz do wnętrza
komórki
Wpuklona błona tworzy w wyniku fuzji
endosom
, który stanowi zamknięte ciałko
otoczone błoną
Po wprowadzeniu do pęcherzyka substancje mogą być przetwarzane w komórce
Egzocytoza:
Przeciwny transport endocytozy
Polega na transporcie obłonionych wakuoli w wnętrza komórki na zewnątrz do
przestrzeni międzykomórkowych
Dochodzi do fuzji cząsteczek i wydalania zawartości poza komórkę
Fuzja umożliwia budowanie nowej błony
Fagocytoza:
Włączanie do komórek dużych stałych cząsteczek (bakterie, fragmenty rozpadłych
komórek) odbywa się poprzez otoczenie wypustkami cząsteczki zabsorbowanej
wcześniej na powierzchni komórki
Tworzy się wakuola (której średnica wynosi powyżej 250 nanometrów), która zawiera
pobrana cząsteczkę
Dołączają się lizosomy, tworząc
fagolizosomy
Pinocytoza:
Transportowanie przez pęcherzyki pinocytarne substancji rozpuszczonych w płynie
otaczającym komórkę
Pęcherzyku te są transportowane we wnętrzu komórki (ich średnica wynosi od 150 –
250 nanometrów)
Transcytoza:
Transportowane są cząsteczki prze cytoplazmę z jednej powierzchni komórki na drugą
powierzchnię
Połączenie endocytozy z egzocytozą
Endocytosomy
wprowadzają materiał do
endosomów wczesnych
, następnie do
endosomów późnych (lizosomalnych)
transportowanych za pośrednictwem
pęcherzyków transportujących
Fotocytoza:
Transport małych cząsteczek przez wytwarzanie wgłówień (jamek/wpukleń) błony,
które zawierają receptory do transportowanych cząsteczek
Endocytoza adsorpcyjna:
Odmiana endocytozy

18
Dwuetapowa
Substancje włączane do komórki są adsorbowane na wewnętrzną powierzchnię
komówki
Tworzy się dołek pokryty
klatryną
(białko)
Dołek się pogłębia i traci klatrynę
Wytworzony pęcherzyk zlewa się z endosomem wczesnym
Heterofagosom
- transportowany materiał zawiera obce bakterie
Autofagosom
– transportowane są zuzyte struktury, własne elementy
Heterofagosomy
i
autofagosomy
mogą się zlewać z lizosomami tworząc
heterofagolizosomy
i
autofagolizosomy
POŁĄCZENIA MIĘDZYKOMÓRKOWE
Funkcjonowanie całych organów możliwe jest dzięki funkcjonowaniu narządów, a te z
kolei dzięki funkcjonowaniu tkanek, a te dzięki funkcjonowaniu komórek
Współpracujące komórki ściśle do siebie przylegają
W miejscach tworzą się specjalne struktury, łączące je ze sobą połączeniami
międzykomórkowymi
Umożliwiają rozpoznanie i przyleganie komórek do siebie i do błony podstawowej
Umożliwiają wzajemną komunikację
Podział:
1. Połączenia zamykające/ścisłe typu
occludens
Bardzo szczelne
Nazywa się je nieprzepuszczalnymi
Reprezentowane przez obwódkę zamykającą
zonula occludens
Szczelne połączenia pomiędzy sąsiednimi komórkami nabłonkowymi, które
biegną wzdłuż obwodu komórki
Nieprzepuszczalne dla jonów i cząsteczek
Ich szczelność wynika z częściowej fuzji błon komórek sąsiednich,
wspomaganą przez białka błonowe (integralne), które tworzą rodzaj sznurów
równoległych do powierzchni nabłonków
Znajdują się w wierzchołkowych częściach komórek nabłonkowych i biegna
wzdłuż bocznych ścian komórki
Oddzielają środowisko zewnętrzne związane ze szczytową częścią komórki od
przestrzeni międzykomórkowej bocznej i podstawy
Są rozwinięte w nabłonkach jelita cienkiego
Zapobiegają przenikaniu enzymów trawiennych ze światła jelita do przestrzeni
międzykomórkowych (dyfuzja wsteczna)
Ograniczają wyspecjalizowany w wchłanianiu obszar nabłonków
Znajdują się w szczelnych połączeniach pęcherza moczowego
Zapobiegają ucieczce wody
2. Połączenia zbierające/przylegania typu
adherens
Zapewniają mechaniczną wytrzymałość
Odporne na rozrywanie
Powstają wskutek połączeń cytoszkieletów sąsiadujących komórek
Tworzą się pomiędzy szkieletami a błoną podstawną
Występują w trzech postaciach:
Obwódka zbierająca
(zanula adherentes)

19
o
Zespala sieć filamentów aktywnych z kadhedrynami błony (cząsteczki
glikoprotein) dwóch sąsiadujących ze sobą komórek nabłonka
o
Znajdują się wdłuż bocznych powierzchni części wierzchołkowych, ale
poniżej połączeń zamykających
Desmosomy (plamki przylegania)
o
Błony przylegających komórek łączą się na ograniczonych
powierzchniach odgrywających rolę nitów
o
W przestrzeni między błonami znajduje się
desmogleina
, która łączy
błony przylegających komórek przy pomocy
krążka
zbudowanego z
desmoplakinowego białka
o
Do niego przylegają
tonofilamenty
(pośrednie elementy
cytoszkieletu)
o
Biegną przez cytoplazmę ku
desmosomom
przeciwległych ścian
komórkowych i wspólnie z
desmogleiną
tworzą pasmo włókienkowe,
które wzmacniają
Hemidesmosomy
o
Łączą komórki nabłonkowe z blaszką podstawną przylegających
komórek
o
Składają się z krążka zagęszczonej cytoplazmy i przy pomocy sieci
filamentów pośrednich łączą się z filamentami z wnętrzem komórki
o
Występują w komórkach wyściełających jelito cienkie, gdzie tworzą
pasmo kwasochłonne
o
Występują w nabłonkach poddawanych dużej sile mechanicznej
(naskórek, nabłonek pochwy, szyjki macicy)
o
Występuje pomiędzy komórkami mięśnia sercowego
3. Połączenia komunikacyjne/jonowo-metaboliczne typu
neksus
Najczęściej występujące połączenia występujące pomiędzy komórkami
Przenikają przez nie jony lub związki drobnocząsteczkowe, mające masę
cząsteczkową do 1500 i rozpuszczalne w wodzie
Służą do komunikowania się między cząsteczkami
Mają kształt pasm biegnących wzdłuż bocznych ścian komórkowych
Każde połączenie przypomina wysepkę poprzebijaną otworkami –
koneksonami
Każdy
konekson
zbudowany jest z sześciu podjednostek białka
transbłonowego –
koneksyny
, leżącego błonach komórek sąsiadujących
W środkowej części
koneksyny
znajduje się kanał o średnicy 1.5 nanometra,
który może się zamykać i otwierać w wyniku zmiany konformacji cząsteczki
koneksyny
W mikroskopie elektronowym widoczne są jako płaski obszar bliskiego
kontaktu, gdzie
koneksony
występują jako ciemne kropeczki
Transport przez ten typ połączeń wymaga energii w postaci ATP i zależy od
stężenia jonu wapnia i wodoru w cytozolu (wzrost ich stężenia hamuje
transport
Występują w komórkach aktywnych elektrycznie (komórki nerwowe; tworzą
synapsę), warunkując przepływ impulsów elektrycznych
Występują również w komórkach mięśniowych i gruczołowych, komórkach
mięśnia sercowego, gdzie synchronizują skurcz i biorą udział ze wzbudzaniu
automatycznych skurczach serca
08.11.2007

20
Specjalizacja powierzchni komórek nabłonkowych
Powierzchnia komórek nabłonkowych jest przystosowana do wielu funkcji –
wchłanianie, transport, oddzielanie od siebie różnych środowisk, a odbywa się to
wszystko dzięki wyspecjalizowanym strukturą na powierzchni komórki
Mikrokosmki:
Są to palczaste wypustki szczytowych części cytoplazmy komórek nabłonków
absorpcyjnych, gdzie tworzą
rąbek szczoteczkowy
(np. w nabłonkach kanalików
nerkowych) lub
rąbek wchłaniający
(np. w komórkach nabłonka jelita cienkiego)
Długość waha się w granicy około 0.5 – 1 mikrometra
Szerokość około 0.1 mikrometra
Na powierzchni jednej komórki może znajdować się do 3000 mikrokosmków
Zwiększają powierzchnię wchłaniania
Pojedynczy mikrokosmek otoczony jest przez błonę komórek jego rdzeni
Zbudowane z 30 równoległych do siebie i do długości osi mikrofilamentów
aktynowych
Na szczycie znajduję się cienka warstwa glikokaliksu
Mogą wykonywać ograniczone ruchy zgodnie z ruchem ślizgowym
Stereocylia:
Długie mikrokosmki
Występują na powierzchniach komórek nabłonka wyścielającego przewód najądrzy
oraz komórek zmysłowych przedsionka i ślimaka (ucho wewnętrzne)
Rzęski i wici:
Mają do siebie podobna budowę
Rzęski występują w dużych ilościach
Nitki występują pojedynczo
Rzęski
Ruchome, nitkowate wypustki, niektórych komórek nabłonkowych (jajowód,
tchawica)
Powodują przesuwanie substancji
Umożliwiają ruch
Ich długość wynosi około 5 – 10 mikrometrów
Szerokość wynosi około 0 – 0.2 mikrometrów
U człowieka ogólna powierzchnia urzęsienia wynosi 0.6 m
2
Witki
Długość około 80 mikrometrów
Występują w plemnikach – nadając ruch
Ich budowa jest podobna w przekroju poprzecznym do budowy rzęsek
Składają się z
aksonemy
, które łączą się z cytoplazmą za pomocą
kinetosomu
(ciałko podstawowe)
Aksonema
zbudowana jest z 9 par częściowo połączonych
mikrotobuli
zlokalizowanych na obwodzie
Składają się promieniście tworząc walec, w środku którego znajdują się 2
niepołączone
Całość tworzy układ 9+2

21
Witki obwodowe połączone są
neksyną
– białko (połączone każda para ze
sobą)
Każda para ma
ramiona dyneinowe
(
dyneina
ma aktywność białka ATP-azy)
Aksonema
zagłębia się kinetosomem w cytpolaźmie, mającym trójkę
mikrotobul (9+3)
Ruch
Odbywa się według mechanizmu ślizgowego
Dyneina
(białko motorowe) ślizga się wzdłuż sąsiednich par mikrotubul
Sąsiednie pary są powiązane
neksyną
, a podstawa jest w ciałku
podstawowym
Ruch ślizgowy powoduje zginanie się rzęski lub witki
20 ruchów na minutę z prędkością 50 mikrometrów/sekundę
Ruch dwufazowy
– niesymetryczny ruch, dwuetapowy
o 1 etap – faza zginania (efektywna)
o 2 etap – faza prostowania (powrotna)
Wici mają ruch śrubowy
Występowanie rzęsek:
Wyraźne w nabłonku dróg oddechowych, gdy przesuwają się po powierzchni komórek
W komórkach nabłonka jajowodu, gdy przemieszczają komórkę uwodnioną w
kierunku macicy
Funkcje barierowe nabłonków:
Tworzą barierę, co wiąże się ze specjalizacja strukturalną
Połączenia zamykające zapobiegają dyfuzji cząsteczek pomiędzy komórkami
(szczytową a podstawową powierzchnią komórki)
Zapobiegają dyfuzji wstecznej
Błona komórkowa u szczytowej części komórki
W drogach moczowych jest duży procent sfingolipidów, które zapobiegają
przechodzeniu płynów – elektrolitów
Desmosomalne i hemidesmosomalne komórki zapewniają mocne mechaniczne
połączenia pomiędzy komórkami i w błonie podstawowej, co zapobiega rozdzielaniu
tych warstw –
bariera mechaniczna
Jądro komórkowe – budowa i funkcje
Jest największym obronionym organellom
Zawiera około 99 % materiału genetycznego
Zostało odkryte pod koniec XVII wieku – 1831 Robert Brown nazwał jądro
nucleusem
Znajduje się tylko u Eukariontów
Funkcje:
Replikacja (powielanie DNA)
Transkrypcja (synteza RNA)
Wielkość i kształt:
Zależy od stanu czynnościowego komórki i typu komórki
Młode komórki intensywnie dzielące się o dużej aktywności metabolicznej

22
Duże jądra
Wyraźne jąderko
Rozproszona chromatyna
Dojrzałe komórki
mniej regularny kształt
Stare komórki
mała aktywność metaboliczna, stosunek powierzchni jądra jest mniejszy do
cytoplazmy
Degenerujące komórki
jądro komórkowe ulega fragmentacji
W fazie G1 są niewielkich rozmiarów
W fazie G2 zwiększają swoje rozmiary
Średnia wynosi około 5 mikrometrów (plemniki) do 40 mikrometrów (w komórkach
jajowych)
Występują u wszystkich eukariontów z wyjątkiem u dojrzałych erytrocytów (kształt
dwuwklęsłego biszkoptu) i w komórkach warstwy rogowej naskórka
Liczba jąder:
Najczęściej występuje jedno jądro
Dwa jądra – hepatocyty (komórki wątroby, nadnercza, komórki gruczołowe żołądka,
pęcherza moczowego)
Wiele jąder – osteoklasty (komórki kostne), chondrocyty (komórki chrzęstne),
komórki mięśni szkieletowych
Lokalizacja:
Najczęściej znajdują się w centrum
Ekscentryczne – przy biegunie
Boczne – pod plazmolemmą (komórki żółte, tłuszczowe, mięśniowe)
Podczas procesów zachodzących w komórce, lokalizacja może ulec zmianie
Składniki jądra:
Otoczka jądrowa
Macierz jądrowa
Chromatyna (skondensowana – heterochromatyna, luźna – euchromatyna)
Jąderko
Perychromatyna
Interchromatyna
Ciałka jądrowe (wtręty jądrowe)
Otoczka jądrowa:
Dynamiczna i asymetryczna struktura
Oddziela zawartość nukleoplazmy jądra od zawartości cytoplazmy komórki
Transport i wymiany zachodzą w ściśle określonych miejscach, przez pory
Wyróżniamy w jej obrębie
Zewnętrzną błonę jądrową (cytoplazmatyczną)
Wewnętrzna błonę jądrową
Przestrzeń perynuklearną (pomiędzy błonami)
Pory jądrowe
blaszka jądrowa (
lamina
), przylegająca do wewnętrznej powierzchni błony
Każda z tych błon jest asymetryczna czynnościowo i strukturalnie

23
Na błonie zewnętrznej znajdują się rybosomy, łączące się z siateczką
śródplazmatyczną
Wyróżniamy różnorodny skład białkowy tych błon
Każda ma typową budowę z dwiema warstwami
Grubość wynosi około 5-8 nanometrów
Szerokość perynuklearnej przestrzeni wynosi około 30 nanometrów
Nie ma ciągłej struktury, ale zawiera liczne otwory (pory)
Bariera między jadrem a cytoplazmą
Aktywnie uczestniczy w transporcie RNA do cytoplazmy
Transportuje z cytoplazmy do jądra białka strukturalne i enzymatyczne
Pory – jądrowy kompleks porowy:
Średnica około 120-150 nanometrów
Ich liczba jest różna i zależy od wieku, aktywności i typu komórki
Przeciętnie bywa około 10-20 porów na mm
3
Budowa:
Jest to złożona cylindryczna struktura białkowa o kształcie oktagonalnym (8
boków złożonych)
Posiada 3 współosiowo ułożone pierścienie (od blaszki zewnętrzny pierścień
cytoplazmatyczny, drugi od strony nukleoplazmy)
3 pierścienie - kompleks 8 wpuklających się promieniście kanałów cytoplazmy
Zlokalizowano tu ponad 100 białek
Nukleoporyny
– rola w transporcie
Blaszka jądrowa (lamina)
Średnia grubość wynosi około 10-100 nanometrów
Przylega do nukleoplazmatycznej powierzchni otoczki jądrowej
Zbudowana jest z sieci włókienek białkowych (laminy) (stanowią 20-25 % wszystkich
białek)
Do wewnętrznej błony otoczki przylegają białka globularne
Funkcje:
Wewnętrzna powierzchnia stabilizuje włókienka chromatyny (łączy się z
fragmentami interfazowych chromosomów)
Laminy biorą udział w procesie fragmentezy i odbudowaniu otoczki podczas
podziałów
Pory w otoczce umożliwiają transport
Każda otoczka składa się z dwóch błon oddzielonych przestrzenią – dyfuzja
jest tu utrudniona
Istnieją jednak pory pomiędzy karioplazmą a cytoplazmą
Małe cząsteczki o masie cząsteczkowej do 40 tysięcy są transportowane bez
trudu
Większe cząsteczki o masie cząsteczkowej ponad 60 tysięcy są transportowane
z trudem, odkształcają się
Cząsteczki są transportowane w obu kierunkach, takie jak białka histonowe,
kompleksy enzymatyczne, duże cząsteczki i podjednostki rybosomów
Chromatyna:
Wybarwia się barwnikami zasadowymi
Zbudowana jest z czterech elementów
Dwóch rodzajów kwasów nukleinowych (DNA i RNA)

24
Białek histonowych
Białek niehistonowych
Białka histonowe
Zasadowe
Maja niską masę cząsteczkową – 10-23 kilodaltonów
Zawierają dużo aminokwasów zasadowych takich jak lizyna, arginina
Jest 5 klas histonów
o H
2
A
o H
2
B rdzenne
o H
3
histony
o H
4
o H
1
łącznik pomiędzy nukleosomami
Nie wykazują swoistości gatunkowej ani tkankowej
Najłatwiej można wyizolować H
1
co świadczy o jego innej funkcji
Interfazowa chromatyna:
Euchromatyna – rozproszona, luźna, w mikroskopie elektronowym tworzy
elektronowo przejrzysty obraz, aktywna transkrypcyjnie
Heterochromatyna – zbita, w mikroskopie elektronowym tworzy gęste
obrazy, często przylega do błony, forma skondensowana, nieaktywna
transkrypcyjnie
Cząsteczki DNA w jednym diploidalnym jądrze osiągają 2 metry długości
Po syntezie jest podwojona ilość – 4 metry
Liczba nukleotydów w chromatynie 2n wynosi 10
9
-10
10
par nukleotydów
Długośc nici DNA po replikacji wynosi 4 metry, więc musi być silnie upakowana, co
jest możliwe Dzięki za wszystko białkom pistonowym
Budowa interfazowej chromatyny:
Nukleonom – 10/6 nanometra
Średnio jest 1,5 * 10
7
nukleosomów w jednym jądrze
Nukleonom składa się z rdzenia – dwa tetrametry białek pistonowych:
o H
2
A
o H
2
B
o H
3
o H
4
140 par zasad jest nawiniętych na każdy rdzeń
Pakowanie:
1 etap upakowania
Tworzenie się nukleofilamentu (z nukleosomów)
Następuje siedmiokrotne skrócenie
Rozluźnianie i zagęszczanie odbywa się Dzięki za wszystko H
1
(struktury
nukleofilamentów)
2 etap upakowania
Dalsze skrócenie odbywa się za pomocą solenoidów – spiralnie zwinięty
nukleofilament
3 etap upakowania
Solenoid tworzy wybrzuszenia w każdej pętli
Długość każdej pętli wynosi 70 mikrometrów
Najwyższy stopień upakowania wynosi 1:10 tysięcy w chromosomach mitotycznych
Dzięki takiemu upakowaniu możliwe jest przekazywanie materiału genetycznego

25
Chromatyna plemników:
Bardzo zbita
Nie ma nukleosomów ani nukleofilamentów
Zginają się, układają koło siebie i łączą za pomocą białek
Chromosomy mitotyczne:
Skondensowana postać chromatyny przygotowana do rozdzielenia i rozdzielenia
komórkom potomnym
Nie ulega transkrypcji i translacji
Kondensacja następuje podczas metafazy, a mniej skondensowana podczas profazy
Liczba i rodzaje ludzkich chromosomów:
Diploidalne jądra komórki somatycznej – 46 chromosomów mitotycznych, które
występują w parach jako chromosomy homologiczne (jeden od ojca, drugi od matki)
Haploidalne – 23 chromosomy
Zestaw chromosomów – 22 par chromosomów (
autosomów
) i jednej pary
heterosomów (chromosomów płciowych)
Poliploidy – wielokrotność haploidów
Tetra- 4*
Penta- 5*
Heksa- 6*
Aneplaudia – liczba różniąca się od wielokrotności liczb haploidalnych
Kształt i budowa chromosomów mitotycznych:
Zbudowane z takich samych składników jak chromatyna
Kilka włókienek spiralnie zwiniętych tworzy chromatynę
Dwie chromatydy połączone w miejscu zwanym przewężeniem pierwotnym
(centromerem)
Zewnętrzną część centromeru otacza kinetochor
Przewężenie dzieli chromosom na dwie półowki
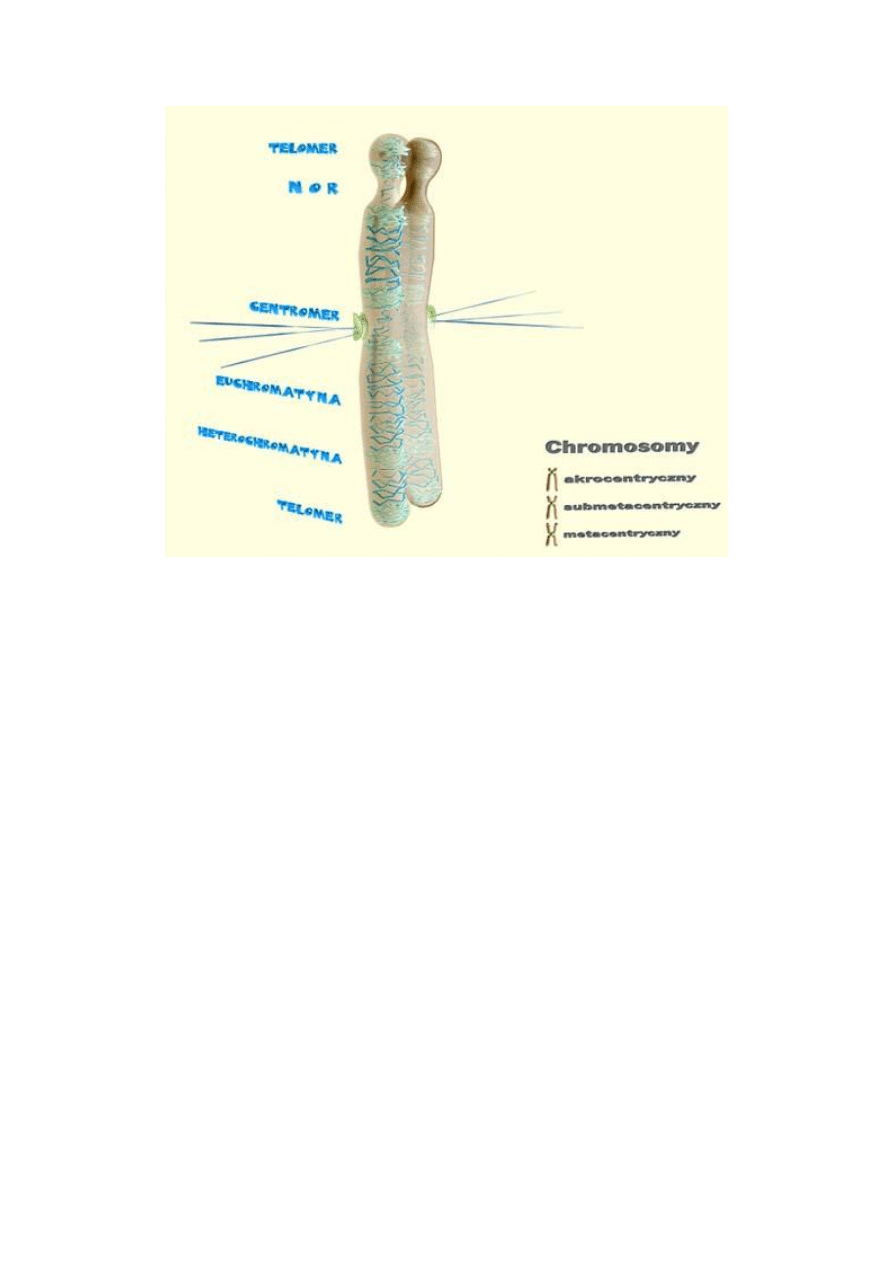
26
Metacentryczne – obie połówki są równe
Submetacentryczne – górna połówka jest dłuższa, dolna krótsza
Akrocentryczne – dolna połówka jest dłuższa, górna krótsza
W pobliżu niektórych końców znajduje się wtórne przewężenie (na 13, 14, 15, 21, 22
chromosomie) – organizuje jąderko
Powyżej wtórnego przewężenia znajduje się satelita (trabant)
Na końcach znajdują się telomery – małe końcowe fragmenty chromosomów,
składające się z DNA nie zawierającego nukleotydów (sekwencja TTAGGG)
Końce telomerów umożliwiają stabilizację struktury
Podczas replikacji zapobiegają przecinaniu przez nukleazy
Telomeraza – wydłużone i nieaktywne u ludzi, po każdym podziale się skraca – jedna
z przyczyn starzenia się komórek
Znajdują się komórki gdzie telomeraza jest aktywna – komórki nowotworowe (powód
nieograniczonego podziału – nieśmiertelne)
15.11.2007
Najsilniej skondensowana chromatyna jest w postaci chromosomów mitotycznych
Materiałem wyjściowym jest włókno chromatynowe wielkości 30 nanometrów
Najbardziej skondensowane są chromosomy metafazowe
Składają się z dwóch ściśle zespolonych chromatyd
Dwa ramiona różnej długości połączone są centromerem – organizator
jąderkowy
Powyżej znajduje się satelita
A na końcu chromatydy znajdują się telomery
Centromer:

27
Jest miejscem, do którego dołączają białka kinetochoru łączące mikrotubule
wrzeciona podziałowego
Kontrolują segregację chromatydy podczas mitozy i mejozy
Molekularna budowa jest różna
Najmniej skomplikowaną budowę mają drożdże
Zbudowany z około 160-220 par zasad
U Drosophila zbudowany jest z około 400 par zasad
U ssaków zbudowany z 4 milionów par zasad
Rejon centromeru łatwo dostrzegalny w mikroskopie elektronowym, gdyż w tym
miejscu ramiona chromosomu są znacznie przewężone oraz dlatego że nie barwi się
barwnikami zasadowymi (jaśniejsze miejsce chromosomu)
W rejonie centromeru organizacja chromatyny jest inna niż w pozostałych odcinkach
Osłonięty specjalnymi białkami, które zabezpieczają na pewnej długości 200-250 par
zasad
Miejsce specyficzne
Kinetochor:
Usytuowany na poziomie przewężenia pierwotnego
Należy do dwóch siostrzanych chromatyd
Kontroluje segregacje chromatyd i depolaryzacje mikrotubul podczas profazy
Inicjuje polimeryzacje i depolaryzacje mikrotubul wrzeciona – warunek ruchu
wrzeciona kariotycznego i segregacji materiału biologicznego
Zakotwiczony do włokien wrzeciona, które skąłda się z 15 mikrotubul
Są strukturami odpowiedzialnymi za ruch chromosomów do przeciwległych biegunów
oraz za precyzyjną orientację chromosomów podczas metafazy w płaszczyźnie na
płytce metafazowej
Budowa:
Składa się z 3 płytek
Zewnętrzna zbudowana z luźnej sieci włókien i filamentów – korona
kinetochoru
Do niej przyczepiają się mikrotobule wrzeciona podziałowego
Środkowa płytka ma włóknistą budowę
Wewnętrzna płytka przylega bezpośrednio do centromerowego rejonu
Ziarnista budowa
Telomery:
Wyspecjalizowane odcinki nici matrycowej DNA, zlokalizowane na każdym z dwóch
końców chromosomów
Składa się z krótkich powtarzających się sekwencji para zasadowych bogatych w
guaninę i tyminę
Przy końcach cząsteczki 3
’
TTAGGG – są n razy powtórzone
Są rozpoznawalne prze telomerazę – enzym komplementarny do sekwencji
telomerowych, służy do syntezy
Odgrywają kluczową role w stabilizacji chromosomów
Są odpowiedzialne za replikację części chromosomów
Chromatydy pozbawione telomerów są niestabilne, łatwo pękają, zlepiają się końcami
z innymi chromatydami
W komórkach rozrodczych i nowotworowych telomeraza jest czynna, więc nie
dochodzi do skracania telomerów, czyli nie dochodzi do śmierci komórki

28
Chromosomy nietypowe/olbrzymie
Duże rozmiary dochodzące do długości 1mm np. w oocytach płazów
W 1889 roku Balbini opisał je w gruczołach ścianach larw owadów
Dzielimy je na dwie kategorie
Poligeniczne – wielowiciowe
o
Występują w gruczołach ślinowych larw owadów
o
Powstają w wyniku wielu endoreplikacji chromatyd, po których nie następuje
podział mitotyczny
o W pojedynczym chromosomie liczba chromatyd zależy od liczby
endoreplikacji
o
W komórkach gruczołów ślinowych zachodzi nawet do 10 endoreplikacji –
DNA jest syntetyzowane, ale nie zachodzi rozdział (1024 chromatyd w jednym
chromosomie)
o
Grubość chromosomów – chromosomy homologiczne są połączone i tworzą
diplosomy, co sprawia wrażenie, że komórki gruczołu ślinowego są
haploidalne (jeden zespół chromosomów)
o
Cechuje je prążkowana struktura i występowanie jednego lub kilku
pierścieniowatych zgrubień –
pierścienie Balbiniego
o
Pierścienie Balbiniego – są to uaktywnione prążki powstałe w wyniku
regularnego ułożenia obok siebie chromomerów (w normalnych
chromosomach występują obok siebie w postaci pojedynczych ziaren) (w
przypadku 1000 ziaren przybierają postać prążków)
o
Każdy z pierścieni składa się od 5-10 prążków
o
W obrębie pierścienia następuje dekondensacje pojedynczych chromatyd
wystających w kształcie pętli poza chromosom
o
Na pętlach zachodzi transkrypcja mRNA, tRNA, rRNA
o
Układ i liczba pierścieni zmienia się regularnie z rozwojem larwy
o
Podczas rozwoju organizmu niektóre geny są zakodowane, a niektóre ulegają
aktywacji
o
Prążki są poprzedzielane fragmentami nie barwiącymi – rejony między
prążkowe składają się z odcinków chromatyny, które uległy kondensacji, mogą
reprezentować sekwencję regulatorową
Szczoteczkowe
o
Przypominają szczotki do czyszczenia butelek od mleka i lamp naftowych
o
Zostały opisane pod koniec XIX wieku – 1882 roku przez Ruckert’a i 1907
przez Marshal’a
o
Występują w oocytach prawie wszystkich kręgowców, w ściśle określonych
stadiach – I profaza mejotyczna - diploten
o
W stadium diplotenu dochodzi do przejściowej dekondensacji chromosomu i
wytworzenia pętli chromatynowych – rozciągają się na błonie chromosomu
przybierając kształt szczoteczek
o Czyli dochodzi do uaktywnienia genów w białkach rybosomalnych, histonów
i żółtkach
o
Jest w nich aktywne 2% DNA potrzebne do rozwoju przyszłego zarodka
o
Stadium diplotenu może trwać bardzo długo
o
Pierwsze zahamowanie w rozwoju mejotycznym powoduje, że kobiety
decydujące się na późną ciąże mogą mieć pewne komplikacje
o
U płazów może trwać nawet rok
o
U człowieka ponad 50 lat

29
o
Rozwija się w piątym miesiącu życia płodowego i jest zahamowany przez
okres dzieciństwa i dojrzewania, aż do pierwszego miesiączkowania
Chromatyna płciowa:
Silnie zasadochłonna grudka chromatyny
Ma średnice około 1 mikrometra
Leży blisko otoczki jąder interfazowych
Silnie skondensowana
Nieczynny jeden z dwóch chromosomów X chromatydy kobiety
Aby nastąpiła kondensacja chromosomu X, musi być drugi chromosom X, którego
chromatyna uległa rozproszeniu
W interfazowych komórkach męskich XY nie ma chromatyny płciowej
Unieczynienie jednego chromosomu kobiety zachodzi podczas rozwoju zaradka
Organizm kobiety składa się z mozaikowato ułożonej grupy komórek zawierający
nieczynny jedne z dwóch chromosomów i chromatynę płciową
W praktyce obserwuje się je w 90% komórek somatycznych kobiety
Podobne grudki można zaobserwować w 30% komórek męskich, przypominających
chromatynę płciową
Kariotyp:
Umożliwia organizowanie liczby i struktury chromosomów mitotycznych
Zespół chromosomów jednej komórki somatycznej uszeregowanych wedle długości i
położenia chromosomów
Kariogram:
Graficzny obraz kariotypu
Analiza jest wykorzystywana w badaniach genetycznych
Badaniach zaburzeń w liczbie i budowie chromosomów
Badania nad przyczyna chorób genetycznych i niepłodności
Analiza zajmuje się szukaniem intensywnie dzielących się komórek
Stosuje się do analizy limfocytów krwi obwodowych – pobudza się do podziału
fitochemagutyniną Pcha
Płytki te hoduje się w komórkach metafazowych in vitro
Fotografuje się je, wycina i układa kariotyp
W celu odróżnienia chromosomów za pomocą barwienia, które uwidacznia prażki
chromosomów, najczęściej wybarwia się prążki G (identyfikuje się nagromadzenie
zasad adeniny i tyminy) i R (identyfikuje się nagromadzenie zasad guaniny i
cytozyny)
Każdy chromosom ma inne geny – poszczególne chromosomy będą się różniły
lokalizacja prążków – identyfikacja poszczególnych prążków chromosomów i zmiana
w ich budowie
Macierz jądrowa:
Struktura poza chromatynowa
Powstaje po trawieniu jadra komórkowego buforami, detergentami i nukleazami
Inne nazwy:
Matriks jądrowa
Nukleoszkielet
Szkielet jądrowy
Jest to rodzaj zrębu podtrzymującego składniki jądra

30
Jego rolą jest regulacja syntezy i transkrypcji DNA
Geny są czynne w pętlach chromatyny w pobliżu miejsca nazywanego macierzą DNA
Nieczynne są w odległych miejscach pętli
Zbudowane z filamentów i ziaren zanurzonych w kariolimfie (sok jądrowy)
Filamenty maja średnicę około 3-5 nanometrów
Ziarna maja średnicę około 15-30 nanometrów
Z filamentami połączone są kompleksy –
replisomy
(replikacja DNA)
Te kompleksy białkowe (spliceosomy) biorą udział w transkrypcji
Reguluje transkrypcję genów odpowiedzialnych za swoistość tkankową różnych
białek
W jej skład wchodzi 23 rodzaje białek
Białka strukturalne – tworzą filamenty
Białka funkcjonalne – enzymy katalizujące syntezę i transkrypcję DNA,
obróbkę heterogennego DNA
8 rodzajów to
matryny
5 rodzajów to
laminy
Filamenty zbudowane sa z matryn
Ziarna zbudowane są z rybonuleny
Perychromatyna:
Poza chromatynowy składnik jadra
Składa się z włókien i ziaren ryboprotein
Małe wysepki w pobliżu otoczki jądrowej
Włókna są różnej długości, średnio mają około 40 nanometrów
Włókna tworzą grupy heterogenne RNA otoczone przez cząsteczki białek
Ziarna są to kompleksy matrycowego RNA, białka, heterogennego i matrycowego
RNA
Miejsce przejściowego przechowywania i obróbki heterogennego DNA
Interchroamtyna:
Występuje w postaci wysepki w różnych częściach jądra
Składa się z cienkich włokien o średnicy 2 nanometrów oraz z ziaren o średnicy 20
nanometrów
Włókna i ziarna zbudowane są z rybonukleoprotein
Odpowiada za zgrupowanie podjednostek rybosomów – przechowywane i
transportowane przez transporter do cytoplazmy
Ciałka jądrowe – wtręty jądrowe:
Znajdują się w wielu rodzajach komórek
Maja średnico około 1000 nanometrów
Kształt owalny
Składają się z dwóch części:
Otoczki (zewnętrzna część)
Rdzenia zbudowanego z różnej wielkości ziaren znajdującego się w centrum
Mogą powstawać w wyniku reorganizacji jąderek
Znaczenie i funkcje są nieznane
Ich liczba zwiększa się w komórkach intensywnie syntetyzujących białka, w
pobudzonych limfocytach B, w komórkach wątroby starszych ludzi

31
22.11.2007
Jąderko:
Nucleus
Nie obłoniona struktura (jedyne organellom)
Kształt sferyczny – okrągły
Brak błony pozwala na duży dynamizm – zmienia się w cyklu komórki
Zanika z profazie (w miejscu organizatora jąderkowego – przewężenie wtórne north),
odtwarza się z telofazie
Liczba obszarów jąderkowych i miejsce występowania są charakterystyczne
U człowieka znajduje się pięć takich miejsc – na chromosomach 13, 14, 15, 21, 22
(akrocentrycznie)
Składniki:
Chromatyna jąderkowa
o
Zbudowana z elementów włóknistych
o
Zdekondensowana postać tych pięciu chromosomów
o
Końcowe fragmenty pętli wytwarzają obszar jąderkowy – jąderko
o
Chromosomy jąderkotwórcze zawierają north, czyli końcowy fragment
pętli
o Chromatyna zawiera geny do syntezy rRNA, sRNA, tRNA, 5sRNA
o
Geny te istnieją w wielu kopiach, a w chromosomach mitotycznych SA
w satelitach przewężeniach wtórnych
o
Dwa rodzaje białek związane są z włókienkami chromatyny –
nukleolina
,
fibrylaryna
Białka
o
Znajduje się tu wiele rodzajów białek
1. nukleolina – fosforylacja nukleiliny – powoduje zmianę
konformacji, dezintegruje i rozprasza składniki jąderka, w interfazie
nukleolina reguluje transkrypcje poprzez zmianę upakowania
chromatyny
2. białko B23 – bierze udział w transporcie prekursorów rybosomów
do cytoplazmy
3. białko Ag-Nor
4. fibrylaryna – obróbka heterogennego RNA (hnRNA)
5. polimeraza RNA
o nukleolina, białko B23 i fibrylaryna mają 19 aminokwasowe
fragmenty, dzięki którym zakotwiczają się w jąderko, biorą udział w
transporcie prekursorów rybosomów do cytoplazmy
Wyróżniamy również w postaci ziaren o średnicy około 15 nanometrów,
równomiernie rozrzucone lub tworzące zgrubienia grudki, zbudowane z
rybonukleoprotein,
są prekursorami podjednostek rybosomów
Rozmiary jąderek zależą od aktywności biosyntezy białka
Wyróżniamy kilka typów jąderek
W komórkach zwierzęcych:
1. jąderko uformowane w nukleolemmę – gąbczaste
aktywne transkrypcyjnie, których struktury nukleoproteidowe tworzą
luźną gąbczastą strukturę
występuję w większości komórek syntetyzujących jąderkowe RNA
2. jąderko zwarte

32
ciasne
złożone z włókien i ziaren jąderkowych
aktywne transkrypcyjnie
można je zaobserwować w szybko rosnących komórkach
3. jąderko pierścieniowate
ma obwodowe rozmieszczenie składników rybonukleoprotein (RNP)
wykazuje zahamowanie syntezy RNA, ale jest to proces odwracalny (są
też aktywne)
4. jąderko zwarte z segregacją składników
poszczególne składniki RNP tworzą oddzielone od siebie strefy
nieaktywne transkrypcyjnie
5. mikrojąderka – jąderka resztkowe
nieaktywne transkrypcyjnie
najczęściej występuje w starych komórkach
Funkcje:
Wytwarzanie składników rybosomów 40S i 60S
o Najpierw geny chromatyny jąderkowej występują dla podjednostek
rybosomów 18S, 5,8S, 28S i odrębnie dla 5S
o
Każdy gen za pomocą polimerazy RNA wytwarza pierwotny transkrypt
RNA (45SRNA)
o
Składają się z około 13 tysięcy nukleotydów
o
Transkrypt ten jest cięty przez nukleazy na te trzy elementy (28S, 18S,
5,8S), które są podjednostkami rybosomów
o
Te podjednostki w jądrze ulegają dalszej reorganizacji i za pomocą
nukleoliny
i
białka B23
są transportowane do cytoplazmy, gdzie są
składane w małe podjednostki 40S i duże 60S i są nalizane na mRNA
Przejściowe gromadzenie matrycowego RNA i syntetyzowanego wzorca DNA
Biosynteza białek
Typy morfologiczne jąderek w komórkach roślinnych:
1. zwarte
o
przewaga składnika ziarnistego na obwodzie, a włóknistego w centrum
o
występuje w większości komórkach roślinnych
2. pierścieniowate
o
oba składniki są zepchnięte do obwodowej części jąderka – powstaje
pierścień
o
występują w komórkach o zahamowanej aktywności syntetycznej np. w
komórkach tkankowych (w hodowlach in vitro), w komórkach
glodzonych
poszczególny typ związany jest ze zmianami sezonowymi
poza tym w profazie mejotycznej (w zygotenie, pachytenie) występuje jeszcze
jeden typ:
3. soczewkowate
o
przylegają do otoczki jądrowej
o
wyraźna segregacja materiału włóknistego i ziarnistego
Nukleogeneza – proces tworzenia jąderka w nowopowstałych komórkach potomnych
1. pierwszy etap
okres ten poprzedzony jest dezorganizacją jąderka macierzystego
w szybko dzielących się komórkach merystematycznych i inicjalnych czapeczki
korzenia już przed profaza zachodzi dezintegracja

33
w wolno dzielących się komórkach zanika pod koniec profazy (w telofazie)
2. drugi etap
odtwarzanie jąderek w merystemie korzeniowym w większości roślin następuje w
późnej telofazie lub na początku interfazy (wczesna faza G
1
)
U roślin istnieją 4 wzorce zachowania się jąderek w mitozie:
1. autonomiczny
jąderko dzieli się z jądrem na dwie połówki
2. persistent
jąderko rozpada się ba nitkowate struktury, które na biegunach komórki tworzą
jąderka potomne
3. semipersistent
jąderko częściowo się zachowuje, a częściowo ulega eliminacji
4. rozproszenie
charakterystyczne dla roślin wyższych
w miarę tworzenia się profazy zachodzi segregacja materiału jąderkowego
glanulazy zanikają
fibrylaza wędruje wraz z chromosomami
Funkcje jądra komórkowego i ich regulacja:
1. replikacja
replikacja DNA całego genomu prowadzi do podwojenia ilości DNA z 2c DNA do
4c DNA i chromosomów z 2n do 4n
proces semikonserwatywny
– zostaje skopiowany każdy z dwóch
komplementarnych łańcuchów cząsteczek macierzystych, w rezultacie czego
powstają dwie nowe cząsteczki identyczne z macierzystymi
zachodzi w fazie S cyklu komórkowego przy udziale kompleksu enzymatycznego
– aparat replikacyjny
skład aparatu replikacyjnego:
enzymy rozplątujące helisę DNA
białko stabilizujące pojedyncze jednoniciowe DNA
polimerazy DNA
poza cyklem komórkowym, może się również odbywać endoreplikacja – prowadzi
do przekształcenia 2C do 4C, zachodzi w chromosomach poligenicznych
etapy replikacji:
początek replikacji
o
rozpoczyna się z udziałem helikaz DNA – białka inicjujące
rozpoczęcie replikacji w określonym miejscu – miejsce inicjacji
„ori”
o
określone sekwencje nukleotydów są rozpoznawane przez helikazy,
które inicjują i oddzielają oba łańcuchy podwójnej helisy DNA
o
stabilizacja pojedynczych łańcuchów DNA
drugi etap replikacji
o
białka wiążące jednoniciowe DNA, tworzą w mikroskopie widełki
replikacyjne
5’ syntetyzowany łańcuch
„ori”

34
3’ widełki replikacyjne
o
Każda z pojedynczych nici służy jako matryca wzdłuż której
przesuwa się aparat replikacyjny
o
Widełki replikacyjne przyjmują kształt litery Y
o
W strefie widełek proces replikacji odbywa się dwukierunkowo
przy pomocy polimeraz DNA od końca 3’
o
Nowy łańcuch syntetyzowany jest od końca 5’
o
Kolejność polimeryzacji wyznacza sekwencję nukleotydów
Przebieg replikacji
o
Do rozpoczęcia syntezy potrzebna jest
prymaza
(enzymsyntetyzujący 9-10 nukleotydowy fragment RNA, służący
jako starter)
o Do otworzenia startera polimeraza DNA, zgodnie z zasada
komplementarności zasad, przyłącza nukleotydy w kierunku 5’ do
3’ w przeciwległych łańcuchach widełek DNA
o
Synteza odbywa się na łańcuchu matrycowym w sposób ciagły
o
A łańcuch wiodący nosi nazwę
nici wiodącej
o
Druga nić nazywa się
nicią opóźnioną
o
Obie są syntetyzowane w oddzielny sposób co wpływa na
asymetrię widełek
o
Polimeraza
DNA łączy poszczególne nukleotydy
o Natomiast
ligaza
łaczy poszczególne fragmenty DNA
o
Wydłużanie łańcucha potomnego odbywa się poprzez przesuwanie
się aparatu replikacyjnego wzdłuż nici DNA
2. transkrypcja
proces enzymatycznej syntezy RNA, odbywającej się na matrycy DNA
(mRNA<<DNA)
zachodzi na terenie jądra komórkowego i podlega na przepisaniu sekwencji DNA
na informacyjny mRNA
enzymy czyli polimerazy RNA to na przykład Pol I, Pol II, Pol III i wiele białek
pomocniczych
w wyniku transkrypcji powstają
transkrypty
(długie łańcuchy), które ulegają
modyfikacji w
procesie dojrzewania
etapy transkrypcji:
wiązanie polimerazy przez matryce przy pomocy dodatkowych białek –
czynniki transkrypcyjne
inicjacja startu transkrypcji przylegającej do promotora (obszar startu)
składa się z kilkudziesięciu – 200 nukleotydów o charakterystycznej
sekwencji TATA
w regulacji szybkości biorą udział geny wzmacniające – wzmacniacze, lub
osłabiające – wyciszacze
elongacja – wydłużanie łańcucha, podczas której powstają kolejne
wiązania fosfodiestrowe
zakończenie transkrypcji objawia się sygnałem, w postaci
charakterystycznej sekwencji AAUAAA
łańcuch nowo syntetyzowanego RNA zawierające heterogenne hnRNA
musi ulec obróbce
35
następuje dojrzewanie i
spilcing
, żeby przekształcić hnRNA w matrycowe
RNA
w dojrzewaniu hnRNA, z
pierwotnego transkryptu
usuwane są
entrony
,
a łączone
egzony
same egzony tworzą matrycowe RNA
modyfikacje posttranskrypcyjne umożliwiają wędrowanie RNA z jądra do
cytoplazmy i odczytanie na rybosomach
transkrypt traci albo zwiększa liczbę nukleotydów
proces zachodzi od końca 3’ do 5’
mogą powstawać nowe trójki kodujące inne aminokwasy
uczestniczą tutaj korygujące sekwencję DNA
jest to jeden z mechanizmów replikacji i ekspresji genów
Naprawa DNA:
u organizmów wyższych utworzyło się wiele systemów naprawy DNA, które mają
zapewnić stabilność i integralność genomów
zabezpieczają również bierność replikacji na poziomie jednego błędu na 10
7
par zasad
enzymatyczne mechanizmy korygują uszkodzenie DNA, powstając spontanicznie
przez czynniki endo- i egzogenne
działają podczas replikacji i w procesach między podziałami komórki
błędy są przyjmowane i korygowane przez różne systemy
jak nie są usunięte błędy, wtedy następują utrwalone zmiany struktury DNA i mutacje
mechanizmy naprawy DNA:
1. przez rewersję bezpośredniego uszkodzenia uszkodzenia w jednej
2. wycinanie zasad azotowych nici DNA
3. wycinanie nukleotydów
4. naprawy błędnie sparowanych zasad azotowych
5. rekombinacja – zachodzi kiedy uszkodzenia następują w dwóch niciach,
zachodzi w obecności nieuszkodzonych homologicznych chromosomów,
wycięcie uszkodzonego odcinka i zsyntetyzowanie nowego odcinka,
wymiana całego dwuniciowego odcinka DNA
29.11.2007
Transport jonowo-cytoplazmatyczny
Informacja genetyczna zawarta jest w jądrze i stąd kieruje cytoplazmą
Regulacja aktywności genetycznej odbywa się przy pomocy białek, które wędrują z
cytoplazmy do jądra
Produkty aktywności genów – kwasy rybonukleinowe, są transportowane z jądra do
cytoplazmy
Wymiana pomiędzy jądrem a cytoplazmą i cytoplazmą a jądrem, jest kontrolowana
Czynnikiem ograniczającym jest rozmiar migrujących cząstek
Wyróżniamy 4 rodzaje transportu:
1. aktywny transport - zachodzi, gdy transportowane są jony i małe cząsteczki,
łatwo przenikając przez otoczkę
2. zachodzi, gdy duże cząsteczki przenikają przez pory jądrowe w otoczce i do
tych cząsteczek zaliczamy
albuminy
i
klobuminy
(są to białka);

36
małe cząsteczki takie jak białka o średnicy 5 nanometrów łatwo przenikają,
natomiast duże przenikają wolniej; cząsteczki powyżej 7 nanometrów nie
przedostają się do jądra przez pory
3. w warunkach in vivo istnieje konieczność transportowania dużych białek
endogennych lub hormonów steroidowych i wirusów(powyżej średnicy 7
nanometrów) i dzieje się to w wyniku endocytozy
4. duże cząsteczki (np. resztki jąderek wyrzuconych z jąder komórek
gruczołowych) przedostają się do cytoplazmy wraz z obiema błonami
jądrowymi, które tworzą otoczkę; błony które są przerwane, łączą się, co
prowadzi do zamknięcia otoczki jądrowej i otoczki wewnątrz pęcherzyk
otoczonego błonami
Totipotencja jądra:
Jądro zapłodnionej komórki jajowej ma pełna informację genetyczną determinującą
rozwój całego organizmu (na tym polega totipotencja)
U roślin i zwierząt w wyniku totipotencji dochodzi do odróżnicowania komórek –
podział, a następnie stopniowego różnicowania oraz do regeneracji całych
organizmów
U zwierząt w dojrzałych tkankach następuje zablokowanie totipotencji jądra
Jądra dorosłych tkanek już nie posiadają takiej zdolności totipotencji
U roślin to zablokowanie określonych systemów jest odwracalne
Warunkiem uaktywnienia całego genomu komórki, jest jej wycięcie spod kontroli
czynników regulujących rozwój całego organizmu
Ma to miejsce w przypadku możliwości regeneracji odciętych organów (rozmnażanie
wegetatywne), wyizolowanych tkanek pojedynczych komórek bądź protoplastów
U roślin determinuje się fakt ujawnienia pełnej potencji komórek, nawet
haploidalnych (komórki generatywne >> ziarno pyłku, haploidalna komórka woreczka
zalążkowego)
U zwierząt totipotencje odznaczają się zapłodnieniem jądra – wczesny etap rozwoju,
ale istnieją komórki macierzyste o totipotencjalnych możliwościach różnicowania się
w inne tkanki
Znajdują się w komórkach embrionalnych i we krwi pępowinowej, tkance mięśniowej
(komórki satelitarne), w wątrobie i trzustce, służąc do wytwarzania dojrzałych
komórek
We wczesnym stadium rozwoju zarodka, nazywane są zarodkowymi komórkami
macierzystymi
Współzależność jądra i cytoplazmy:
Czynniki zawarte w cytoplazmie mogą indukować aktywność jądra, które
charakteryzuje się większym stopniem represji (zahamowania) aparatu genetycznego
W warunkach laboratoryjnych powstałych w wyniku fuzji w heterokarionach
(komórki w których we wspólnej cytoplazmie istnieją dwa gatunkowo odmienne
jądra
Występuje zawsze indukcja pozytywna pojawienia się takich właściwości w jadrze,
które charakteryzują jądro drugiego składnika heterokarionu np. indukcja replikacji
DNA lub synteza RNA
Jedno jądro stymuluje, pobudza drugie

37
Fuzje komórek w różnych fazach cuklu mitotycznego np. w komórkach w fazie S z
komórkami w fazie G
1
(faza S jest bardziej zaawansowana), powoduje przyspieszenie
replikacji DNA w komórkach będącej w fazie G
1
Natomiast fuzja komórek w fazie G
2
z komórkami w fazie S, nie następuje replikacja
w jadrze w fazie G
2
, co świadczy o braku kompetencji chromatyny w fazie G
2
wobec
induktorów syntezy w fazie S
Czas trwania fazy G
2
przedłuża się o okres niezbędny do zakończenia replikacji w
jądrze w fazie S
Fuzja komórek w mitozie z komórkami w interfazie (będące w różnym stanie)
prowadzi do przedwczesnej kondensacji chromosomu indukowanej przez komórki
będące w mitozie
Podobnie jak w przypadku indukcji syntezy DNA lub RNA czynniki indukujące
przedwczesną kondensacje chromosomów nie SA specyficzne dla gatunku
Taka zdolność do indukcji replikacji DNA w komórkach nie dzielących się w wyniku
fuzji z dzielącymi się zostało wykorzystane do otrzymywania przeciwciał
monoklinalnych
Trwałą zdolność do
proliferacji
(dzielenia się) wykazują komórki nowotworowe,
które mogą przekazywać to komórkom prawidłowym w wyniku fuzji z prawidłowymi
Sygnały emitowane są przez obcą cytoplazmę i przenikający do jądra drugiego skłąd
heterokarionu (zlane jądro z dwóch różnych komórek) są przypuszczanie białkami
regulatorowymi, które posiadają zdolność derepresji (odblokowywania) określonych
obszarów chromatyn
Mitochondria:
Są obecne we wszystkich komórkach Eukariotycznych poza wyjątkami
Erytrocyty
Ameby patogenne
Stanowią odrębny od cytoplazmy przedział metaboliczny oraz z chloroplastami są
miejscem pozajądrowego genomu (posiadają DNA) i mają zdolność do jego ekspresji
(ujawniania)
Większość białek importuje z cytoplazmy mitochondrialne DNA (mtDNA) i stanowi
0.2 % całkowitego DNA
Otoczone podwójną błoną zewnętrzna i wewnętrzną
Mają kształt walca lub soczewki
Błony maja typową budowę dwuwarstwy białkowo-lipidowej
Błona wewnętrzna jest wpuklina i pofałdowana, tworzy grzebienie różnego kształtu
Blaszkowaty
Rurkowaty
Pomiędzy błonami znajduje się przestrzeń międzybłonowa
Błona wewnętrzna otacza przestrzeń zwaną matriks mitochondrialne
Liczebność i struktura jest różna w poszczególnych typach komórek
W komórkach roślinnych jest małą liczba mitochondriów np. w komórkach
epidermy korzenia są nieliczne, a w komórkach aktywnych metabolicznie
stanowią aż 20% objętości cytoplazmy
W komórkach zwierzęcych w szczególności u kręgowców każda komórka
zawiera kilkaset mitochondriów (komórki wątroby mogą mieć ich nawet do
kilku tysięcy)
Liczba mitochondriów jest precyzowana do intensywności metabolizmu
tlenowego i zapotrzebowania komórki na energię
Stanowią centra energetyczne, elektrownie komórkowe

38
Wielkość jest zróżnicowana
Długość stanowi około 0.5 – 2 mikrometrów
Szerokość stanowi około 0.2 – 0.8 mikrometrów
Wielkość i kształt zmieniają się i zależą o stadium rozwojowego komórki oraz
aktywności metabolicznej
Mitochondria mogą się łączyć (fuzja) lub mogą się rozpadać na mniejsze jednostki
Fuzja występuje u roślin, a rozpad u drożdży
Lokalizacja:
Nie mają stałego miejsca w cytoplazmie
Ich występowanie może mieć związek z zapotrzebowaniem energetycznym
W nabłonkach komórek rzęskowych znajdują się w części podstawnej błony,
przez którą zachodzi transport
W błonie wewnętrznej są trzy rodzaje białek
o
Białka uczestniczące w transporcie metabolitów do i z mitochondriów
o
Białka enzymatyczne łańcucha oddechowego biorące udział w
oksydacji związków
o
Białka kompleksu enzymatycznego zawierające syntetazę ATP w
postaci grzybków 0 nazwane cząsteczkami transportu elektronów
Błona zewnętrzna:
o
Transbłonowe kompleksy białek, uczestniczące w transporcie różnych
związków
o
Białka enzymatyczne do transformacji lipidów oraz monooksygenazy
P-450
W macierzy mitochondrialnej znajduje się:
Wiele enzymów katalizujących reakcję przemian kwasów tłuszczowych i
kwasu pirogronowego (wytwarzany acetylo-CoA utlenia go w cyklu Krebsa w
CO
2
i NADH)
Wolne rybosomy (biosynteza białka)
DNA mitochondrialne
Jony wapnia
Enzymy fosforyzujące nukleotydy
Mitochondria połączone są z cytoszkieletem komórki za pomocą mostków – białko
MAP
2
W błonach mitochondriów znajdują się kanały wapniowe
Kanał porynowy
Kanał potasowy
Kanał chlorkowy
Funkcje:
Następuje w nich uwalnianie i magazynowanie energii w postaci ATP
(„elektrownie”)
W wyniku przemian kwasów tłuszczowych i kwasu pirogronowego z
wytworzeniem acetylo-CoA
Mitoplasty
– mitochondria pozbawione błony zewnętrznej
Submitochondria (SMP
) – powstają po użyciu ultradźwięków, które niszczą rozrywając obie
błony, zewnętrzną niszczą i po jej samozasklepieniu powstają pęcherzyki otaczające błonę
wewnętrzną
Mitochondria jako organelle wytwarzające ATP:

39
ATP służy jako najlepszy pośrednik pomiędzy reakcjami wyzwalającymi a
pochłaniającymi energię - uniwersalny przenośnik energii swobodnej
Podczas hydratazy ATP powstaje dużo energii i ADP, który wykorzystywany jest do
syntezy ATP
Oprócz tego tworzy się ortofosforan P
1
ATP ADP + P
1
+ energia
Ortofosforan w połączeniu z nieaktywna cząsteczką, energetyzuje ją przy pomocy
przechwyconej energii hydrolizy ATP
W ten sposób energia wiązań chemicznych ATP jest wykorzystywana do różnego
rodzaju pracy
Reakcja odwrotna
ADP + P
1
ATP
Może być zredukowana w wyniku fosforylacji oksydacyjnej , które zachodzi w
wewnętrznej błonie mitochondrialnej
Elektrony przenoszone przez łańcuch oddechowy pochodzą z NADH i FADH
2
,
tworzących podczas glikolizy cyklu kwasu cytrynowego i utleniania kwasów
tłuszczowych
Mutacje genomu mitochondrialnego:
Mitochondrialne DNA zbudowane jest z podwójnej kolistej nici pozbawionej
intronów i białek pistonowych
Na przykład u człowieka genom mitochondrialny zawiera 16569 par nukleotydów i
koduje dwa rodzaje rRNA, 22 rodzaje tRNA, 13 polipeptydów, które budują
kompleksy łańcucha oddechowego i synteza ATP
Mutacje mogą być
1. punktowe
2. delecje (ubytki)
prowadzą do zaburzeń syntezy białek mitochondrialnych, transportu jonów
wzdłuż łańcucha oddechowego, upośledzeniem fosforylacji oksydacyjnej i
funkcji energetycznej
w komórkach istnieje tysiąc zmutowanych kopii modna
o wiele mniej kopii zawierających patogenne mutacje
kliniczna ekspresja zaburzeń (choroba mitochondria) zależy od stosunku
mitochondrialnego do prawidłowego DNA – dziecko dziedziczy chorobę tylko
po matce
Powstanie mitochondriów:
Mitochondria powstają w komórkach de novo (od nowa) z podstawowych związków
organicznych
Powstają z różnych innych organelli komórkowych
Wyjścia na drodze podziału istniejących komórek mitochondrialnych
06.12.2007
Chloroplasty jako transformatory energii:

40
Występują w komórkach roślinnych
Zaliczamy je do dużej rodziny plastydów
Występują wyłącznie w komórkach Eukariontów
Mają zdolność samopowielania
Do plastydów zaliczamy 4 grupy:
Proplastydy – grupa wyjściowa
Chloroplasty – umożliwiają fotosynteze, zapewniają samożywność
Chromoplasty
Leukoplasty
Podstawowym produktem fotosyntezy jest glukoza
Chloroplasty znajdują się tuż pod błoną
W jednej komórce ich liczba wynosi od około 20-40
Pod względem wielkości i kształtu najbardziej zróżnicowane są w glonach
Otoczone są podwójną błoną białkowo lipidową
System błon tworzą płaskie woreczki – tylakoidy
Budowa błon chloroplastów:
Błony są typowymi białkowo-lipidowymi błonami
Proporcje lipidów bo białek są różne
Błona wewnętrzna jest przepuszczalna dla jonów i związków drobnocząsteczkowych
Błona zewnętrzna jest wybiórczo przepuszczalna
Lipidy tylakoidów stanowią 35-40 % błon
W tych 35-40 % znajduje się 15 % fosfatydyloglicerol
Błona tylakoidów w miejscach gdzie zachodzi faza świetlna fotosyntezy, zbudowana
jest głównie z galaktolipidów, które stanowią do 75% lipidów
W błonach tylakoidów znajduje się zielony barwnik – chlorofil, jak i również
występują karotenoidy (karoten - barwa pomarańczowa, ksantofil – barwa żółta) oraz
fikogliny (fikoerytryna i fikocyjanina)
Stroma zawiera oprócz ziaren skrobi, plastoglobule, a zwłaszcza chinony, DNA,
rybosomy, enzymy białkowe związane z replikacją DNA, transkrypcją i biosyntezą
białek
Chloroplastowy DNA jest trudno dostrzegalny nawet w mikroskopie elektronowym, a
rybosomy są mniejsze od cytoplazmatycznych
Peroksysomy jako pierwotne utleniacze:
Zwane inaczej
mikrociałami
Występują nielicznie we wszystkich komórkach Eukariotycznych
Są to małe pęcherzyki o średnicy 0.5 – 1.5 mikrona
Otoczone są pojedynczą błoną
Pełnią rolę organelli utleniających, ale nie syntetyzują ATP
Głównym układem enzymatycznym jest system oksydoreduktaz flawinowych i
katalazy, które utleniają substraty, produkują H
2
O
2
(nadtlenek wodoru) i rozkładają go
do wody i ½ O
2
Rozkład H
2
O
2
odbywa się dwoma sposobami
1. z udziałem katalazy
2. z udziałem peroksydazy
Zawierają około 50 rodzajów enzymów
Oksydoreduktazy – metabolizujące nadtlenek wodoru
Enzymy katalizujące rozkład puryn
Enzymy cyklu glioksanolowego (tylko u roślin w glioksysomach)

41
Budowa i funkcje peroksysomów w komórkach zwierzęcych:
Zawierają rdzeń – nukleoid, który jest od zewnątrz otoczony macierzą, która jest
pojedynczą błoną zawierającą około 10 białek
Krystaliczny rdzeń składa się z wielu rurek, które gdy posiadają kilka warstw, tworzą
sześcian, a jeżeli tworzą dwuwarstwe wtedy jest to typ blaszkowy
Peroksysomy zużywają około 20% tlenu (uwolnionego podczas rozkładu nadtlenku
wodoru) do bezpośredniego utleniania różnych związków
Podczas utleniania powstaje energia wydzielana w postaci ciepła
Bierze udział w pomocniczej roli w komórkach wątroby w utlenianiu alkoholu
etylowego
W komórkach mięśnia sercowego odbywa główna rolę w utlenianiu alkoholu
etylowego
Przeprowadzają częściową
β-oksydację
kwasów tłuszczowych – zainicjowaną przez
oksydazę acetyloCoA, a nie jak w przypadku mitochondriów przez dehydrogenazę
Produktem ubocznym jest nadtlenek wodoru
Peroksysomy roślinne:
Nazywane są
glioksysomami
Opisane zostały po raz pierwszy w 1967
Zawierają
liazę izocytrynianową i syntetazę jabłczanową, izocytrynianową,
dehydrogenazę jabłczanową, akonitrazę
Peroksysomy liści współdziałają z chloroplastami i mitochondriami w procesie
oddychania
System GERL:
System błon wewnętrznych, w którego skład wchodzą aparat Golgi’ego,
endoplazmatyczne retikulum, lizosomy i wakuole
Po raz pierwszy został opisany w 1974 roku
System ten obejmuje błony, które wchodzą w skład różnych szlaków transportu
komórek Eukariotycznych
Pomiędzy aparatem Golgi’ego, endoplazmatycznym retikulum, lizosomami i
wakuolami odbywa się proces przepływu przez błony, sukcesywnie tworzących się
pęcherzyków w obrębie jednej struktury, transportowanych i łączenia się z drugą
strukturą w wyniku fuzji
Przykładem mogą być białka sekrecyjne (wydzielające), które zamykają szlak w
retikulum endoplazmatycznym, transportowane są do aparatu Golgi’ego, aż w końcu
znajdują się w wakuolach lub na powierzchni komórki
System bierze udział w powstawaniu organelli – odgrywa kluczową rolę w
gromadzeniu różnych substancji, biosyntezie i transporcie materiałów prowadzonych
do pozaplazmatycznych obszarów komórki
Transportujące pęcherzyki przenoszą białka, lipidy, polisacharydy pomiędzy
organellami do ściany lub błony komórkowej
Poszczególne organelle wyposażone są zestaw błonowych rozpuszczalnych białek,
które są syntetyzowane na szorstkim retikulum endoplazmatycznym i pełnią wiele
funkcji związanych z metabolizmem komórki
Retikulum endoplazmatyczne ER:
Jego ilość i budowa zależy od typu komórek, od stanu fizjologicznego i etapu
rozwojowego komórek

42
Ich duża różnorodność spowodowana jest wielością funkcji jakie spełniają w komórce
Funkcje:
Biosynteza białek i lipidów
Transport białek i błon w obrębie komórek i pomiędzy komórkami
Regulacja warunków jonowych, a zwłaszcza poziomu jonów wapnia i
protonów w cytoplazmie
Gromadzenie białek zapasowych i lipidów
Budowa:
Błony retikulum endoplazmatycznego w komórkach roślinnych tworzą sieć
utworzoną z
tubul
(rurek) – wtedy jest to gładkie retikulum
endoplazmatyczne, jak i
cystern
– wtedy jest to szorstkie retikulum
endoplazmatyczne
W komórkach zwierzęcych jest zbiorem kanałów i cystern zamkniętych błoną
Ziarniste retikulum endoplazmatyczne RER:
Tworzy długie błoniaste kanaliki – cysterny, które od zewnątrz pokryte są ziarnami
rybosomów (nie są stale nimi pokryte, tylko wtedy gdy zachodzi synteza białek –
translacja, natomiast po translacji rybosomy odłączają się od cystern, proces zachodzi
cyklicznie z krótkimi przerwami)
Światło kanalików RER jest miejscem, w którym zachodzą procesy po translacyjne i
przygotowanie białek do dalszej drogi np. do wydzielania z komórek lub wbudowanie
w inne układy
Błona budująca cysterny RER oddziela ich treść od cytoplazmy i łączy się z
zewnętrzną błoną osłonki jądrowej
W wielu typach komórek, gdzie RER jest słabo rozwinięte lub nie ma go wcale, to
jego rolę przejmuje otoczka jądrowa
Dwuwarstwa lipidowa jest bardziej płynna niż błony komórkowej, ponieważ
fosfolipidy
mają duży stopień nasycenia kwasów tłuszczowych, poza tym zawierają
więcej
fosfatydylocholiny
a mniej
cholesterolu
Poza tym warstwy błony mają podobny skład i nie ma asymetrii
Białkowy składnik błony tworzą:
Specyficzne białka integralne – odpowiedzialne za modyfikacje
postranslacyjne i eksport białek
Białka enzymatyczne
o
Związane z potranslacyjną modyfikacją białek eksportowych
o
Związane z syntezą lipidów
o
Związane z utlenianiem i redukcją lipidów
o
Związane z transportem substancji do wnętrza cystern siateczki RER
W błonach RER znajdują się enzymy katalizujące procesy glikolizacji białek i
lipidów, czyli transferazy glikozydowi
Białka transportują cząstki lipidów do błon innych organelli
Są miejscem enzymów utlenionych lipidów
RER jest obecna we wszystkich komórkach Eukariontów, z wyjątkiem erytrocytów i
plemników
Najliczniej występują w komórkach wyspecjalizowanych w biosyntezie białek
eksportowych np. w komórkach pęcherzykowych trzustki (syntetyzują enzymy
trawienne), w plazmocytach (syntetyzują gammaglobuline), w fibroblastach (komórki

43
tkanki łącznej, syntetyzują kolagen), w komórkach wątroby (w zespołach cystern
ułożonych równolegle lub owiniętych wokół mitochondium)
W komórkach roślin wyższych RER występują pod postacią pojedynczych kanalików
lub w zespołach cystern
Może się komunikować przez plazmodesmy (połączenia międzykomórkowe), tworząc
jeden wspólny ciąg (system kanałów RER)
Udział RER w procesie syntezy białek:
Do białek syntezy w błonach na rybosomach RER zaliczamy
1. białka integralne
2. białka lizosomalne
3. białka sekrecyjne/eksportowe/wydzielnicze
Mogą przejść przez kilka barier błon hydrofobowych (błony RER, aparatu
Golgi’ego, błony komórkowe), aby syntetyzowany łańcuch mógł pokonać
barierę RER
Jego pewny odcinek musi być uporządkowany w odpowiednie białka
odpowiedniej sekwencji – sygnał
Gładkie retikulum endoplazmatyczne SER:
Sieć rozgałęzionych kanalików
Występuje we wszystkich komórkach Eukariotycznych z wyjątkiem erytrocytów
ssaków
Odgrywa ważną role w procesie neutralizacji leków i trucizn
Bierze również udział w hydrolizie glikogenu i jednocześnie podnoszenia stężenia
glukozy we krwi
Bierze również udział w syntezie lipidów – trójgliceryny, cholesterolu, steroidów,
fosfolipidów
Enzymy są zlokalizowane w błonach SER komórek gruczołów dokrewnych
produkujących hormony steroidowe (testosteron) i komórkach syntetyzujących
komórki nadnerczy (SER występuje w dużych ilościach)
Odmiana siateczki śródplazmatycznej:
Sarkoplazmatyczna siateczka
Występuje w komórkach mięśni (prążkowanych poprzecznie)
Jest wyspecjalizowaną siateczką gładką
Jej zadaniem jest magazynowanie jonów wapnia, co umożliwia cykliczne
uwalnianie i rozdział jonów wapnia do sarkoplazmy (rozkurcz i skurcz
włókien mięśniowych)
Funkcje retikulum endoplazmatycznego:
W przedziale jądra stanowi przegrody pierwotnej ściany komórkowej
Bierze udział w organizacji wrzeciona kariokinetycznego
Może być źródłem błon do odtwarzania otoczki jądrowej
Kontrolują stężenie jonów wapnia w obrebie strefy podziałowej
Miejsce zakotwiczenia elementów cytoszkieletu
Podczas podziału cytoplazmy wychwytuje i utrzymuje w rejonie tworzenia przegrody
pierwotnej pęcherzyki z diktiosomów aparatu Golgi’ego i umożliwia ich zlewanie i
zapewnia właściwą płaszczyznę przegrody pierwotnej
Bierze udział w tworzeniu się
kalazowych
zgrubień ściany komórkowej
Aparat Golgi’ego:

44
Należy do układu wakuolarnego cytoplazmy z retikulum endoplazatycznym, osłonką
jądrową i z systemem endosomowolizosomalnym
Składa się z wielu płaskich błoniastych cystern (od 6 – 20) ułożonych w stos (jedna
nad drugą) i poza tym rozdętych pęcherzyków na końcach (diktiosomy)
Otaczają go z wszystkich stron pęcherzyki
Występują we wszystkich komórkach Eukariotycznych z wyjątkiem erytrocytów
Jest zlokalizowany w pobliżu jądra i cystern szorstkiej siateczki śródplazmatycznej
Wyróżnia się dwa bieguny bądź sieci:
1. biegun/sieć syntezy cis
Występuje w pobliżu jądra
Znajdują się tu zbiorniki zbudowane z błony z rybosomami
2. biegun/sieć syntezy trans
Występuje w pobliżu błony komórkowej bądź ściany komórkowej
Błony są gładkie
o
Transport z sieci cis do sieci trans odbywa się przy pomocy pęcherzyków
fuzujących z innymi
Wyróżnia się dwie formy aparatu Golgi’ego:
1. siateczkowa – występuje w komórkach kręgowców (wyjątek stanowią oocyty i
plemniki)
2. diktiosomalna – występuje w komórkach roślinnych i u bezkręgowców
Funkcje:
Modyfikowanie struktury makrocząsteczek
Segregacja (grupowanie według budowy chemicznej) makrocząsteczek
Kierowanie transportem makrocząsteczek
Makrocząsteczki białek przechodzą przez aparat Golgi’ego i ulegają w nim
zmianom kowalencyjnym – polegają na modyfikowaniu oligosacharydów oraz
dodawania do cząsteczek białkowych grup siarczanowych i kwasów
tłuszczowych
Zachodzi tu glikolizacja białek i lipidów, proteogliaknów, które powstaja w
kanalikach RER
Zmodyfikowane cząsteczki są grupowane według budowy chemicznej i są
otoczone błoną w sieci trans
A pęcherzyki, które powstają w sieci trans mogą być transportowane w dwóch
kierunkach
1. ku endosomom późnym ,a następnie lizosomom (ulegają strawieniu)
2. ku błonie komórkowej (są na ogół pęcherzykami wydzielniczymi –
typowa egzocytoza)
Rodzaje transportu pęcherzyków wydzielniczych:
1. konstruktywny
Pęcherzyki są wytwarzane przez pączkowanie i transportowane w sposób
ciągły, od sieci trans do błony bez udziału sygnałów z zewnątrz
W ten sposób białka i lipidy dostarczają nowe składniki błonie, a zawartość
pęcherzyki wydzielają na zewnątrz
2. wybiórczy
Regulowany jest przez sygnały z zewnątrz np. wydzielanie hormonów
następuje pod wpływem gruczołów przysadki tarczycy
Bierze tu udział
klatryna
(białko), które wiąże się z powierzchnią błony,
powodując jej wybrzuszenie i powstanie pęcherzyka

45
13.12.2007
Recyrkulacja błony:
W wyniku fuzji pęcherzyków egzocytarnych z błoną, zachodzi wydzielanie zawartości
i wbudowanie w błonę komórki
Te fragmenty błony są dalej wykorzystywane do tworzenia pęcherzyków
endocytarnych
Oba procesy są w równowadze dynamicznej
Takie krążenie elementów błon podczas endocytozy, egzocytozy i pączkowania to
recyrkulacja błony
Roślinny aparat Golgi’ego jest odpowiedzialny za syntezę polisacharydów
wchodzących w skład ściany komórkowej
A zwierzęcy aparat Golgi’ego jest odpowiedzialny za syntezę glikolipidów
tworzących błony komórkowe
Endosomy:
Błoniaste struktury cytoplazmy, wstępujące w postaci zbiorników, cewek i
pęcherzyków
Biorą udział w endocytozie – transporcie cząsteczek przez błony
Endosomy wczesne
Endocytowany materiał pojawia się po kilku minutach
Zlokalizowane są w pobliżu błon komórkowych
Endosomy późne
Materiał endocytów pojawia się po około 20 minutach
Znajdują się dalej od błony komórkowej, w pobliżu jądra
Zawartość endosomów wczesnych i białka błonowego jest przenoszona do
endosomów późnych przy pomocy pęcherzyków transportujących
A z endosomów późnych powstają lizosomy (na skutek przekształcenia)
Lizosomy mogą też powstawać w sieci trans aparatu Golgi’ego
Lizosomy:
Średnica 1 mikrometr
Wnętrze lizosomów ma odczyn kwaśny
Zawiera wiele enzymów hydrolitycznych
Proteazy
Lipazy
Nukleazy
Fosfatazy
Sulfatazy
Glikozydazy
Enzymy hydrolityczne są syntetyzowane w RER, skąd są transportowane do aparatu
Golgi’ego, gdzie ulegają modyfikacji
Funkcje lizosomów i endosomów:
Endocytowany materiał przechodzi do pęcherzyków transportujących a z nich do
endosomów późnych, lub pęcherzyki transportujące przenoszą materiał do
przeciwległej błony komórkowej, gdzie uwalniają swoją zawartość – tzw.
transcytoza
(z jednego końca komórki do drugiego)

46
Biorą udział w trawieniu wewnątrzkomórkowym – cząsteczki lub makrocząsteczki
dostające się do komórek przez fuzję i endocytozę są trawione przez hydrolazy w
endosomach późnych przekształconych w lizosomy
(koniec informacji dotyczących aparatu Golgi’ego)
System GERL
Wakuole:
Występują tylko w komórkach roślinnych
Ich liczba i wymiary zależą i różnią się od rodzaju tkanki i stadium rozwojowego
Komórki merystematyczne mają małe i liczne wakuole (średnica 1 mikrometra)
Komórki dojrzałe, zróżnicowane np. komórki mezofilu mają jedna dużą wakuolę
zajmująca nawet 90% objętości komórki
Komórka może zwierać kilka takich samych wakuoli, lub mogą występować
funkcjonalnie różne wakuole
Wyróżnia się co najmniej dwa typy wakuoli:
1. LV – wakuole lityczne
2. PSV – wakuole gromadzące białka zapasowe
W większości typów komórek oba te typy łączą się ze sobą, ale nie wiele wiadomo o
dalszych etapach rozwoju tych wakuol, a zwłaszcza wakuol LV
Są to najbardziej funkcjonalne organelle w komórce
Pełnią funkcje statyczne – zwiększają powierzchnię w stosunku to objętości
Pełnią funkcje dynamiczne – są zdolne do akumulacji białek, związków chemicznych
i jonów
Pełnią funkcje obronne i wabiące
Błona otaczająca wakuole –
tonoplast
– wyposażony jest w kompleksy białkowe i
kanały jonowe, wodorowe, przenośniki cukrów i innych związków, dzięki za
wszystko czemu jest możliwy transport tych związków do wakuoli
W tonoplaście znajdują się dwie pompy protonowe:
H
+
ATP-aza dostarczają energie do różnych
H
+
pirofosfataza procesów transportu wakuolarnego
Funkcje wakuoli:
Gromadzenie substancji np. barwniki kwiatowe – antocyjaniny, flawonoidy,
czerwone betalainy ( w korzeniach buraka)
Zdolne do przeprowadzenia hydrolizy – zawierają hydrolazy,esterazy,
nukleazy, peroksydazy
Utrzymywanie odpowiedniej wartości turgoru – wnętrze zawiera związki
mineralne, wysokie stężenie jonów nieorganicznych i duże ilości cukrów,
kwasów organicznych, aminokwasów – ujemny potencjał roztworu uzyskuje
wysoką wartość i reguluje turgor komórki
Wakuola – „śmietnik komórki” – ma znaczenie w utrzymywaniu wewnętrznej
homeostazy
Funkcje obronne:
Do pełnienia tych funkcji służą
białka toksyczne
, które są gromadzone w
wakuolach wielu nasion, są to tzw. pektyny, poza tym inhibitory proteaz i inne
toksoalbuminy, które pełnią dwie funkcje:
o
Oprócz roli obronnej stanowią także zapas azotu
Po zranieniu komórki rośliny lub infekcji, ulega nasileniu synteza białek
ochronnych, takich jak hitynaza, inhibitor proteaz, które są przetransportowane
do wakuoli

47
Gromadzone są też morfiny i berberyny w wakuolach, które są toksyczne dla
roślin
Substancje te (wtórne metabolity) uzupełniają funkcje obronne roślin
Niektóre związki są kierowane przeciw jednemu organizmowi, lub wielu
przeciwnikom
Ale związki te mogą zwiększać atrakcyjność dla zwierząt rozsiewających
nasiona i zapylających kwiaty
Np. antocyjaniny są związkami szkodliwymi, ale jednocześnie zwiększają
atrakcyjność i są antybiotykami
Mechanizmy umożliwiające gromadzenie substancji w wakuolach:
Pobieranie substancji przez tonoplast (błona ta nie różni się pod względem
budowy od błony komórkowej) – transport na drodze dyfuzji, przez
przenośniki, kanały i pompy
Transport aktywny – wbrew gradientowi stężenia
Mechanizmy wychwytywania – przyłączanie wtórnych metabolitów do białek,
np. krystalizacja, bądź tworzenie kompleksów alkaloidów z kwasem
mekonowym
Cytoszkielet:
Cytoplazma wszystkich Eukariontów zawiera trójwymiarową sieć włókien
białkowych, łączących składniki cytoplazmy ze sobą i z plazmolemmą (błoną)
Jest t zespół włókien białkowych, decydujący o kształcie komórek i jej wewnętrznej
organizacji
Odpowiada za transport i rozmieszczenie organelli komórkowych oraz za skurcz
komórki
Trzy elementy cytoszkieletu
1. mikrotubule – największa śrenidca
2. filamenty pośrednie
3. mikrofilamenty – najmniejsza średnica
Mikrotubule:
Rurki o średnicy około 20 nanometrów, puste w środku
Grubość ściany około 5 nanometrów
Występują w cytoplazmie pojedynczo lub tworzą układy, np. wrzeciono
kariokinetyczne lub rzęski
Złożone są dwóch białek
1. tubulina α – tworzy biegun „-
„
2. tubulina β – tworzy biegun „+”
Obie tubuliny tworzą heterodimer
W niektórych komórkach znajduje się też tubulina γ, nie wchodząca w skład
mikrotubul, ale tworząca kompleksy z białkami inicjującymi polimeryzację
mikrotubul, które występują w centrosferze wokół centrioli (w komórkach
zwierzęcych są to tzw. centra polimeryzacji mikrotubul)
γ tubulina znajduje się w cytoplazmie w komórkach roślinnych
Dimery tubuliny α i β łatwo polimeryzują tworząc
protofilamenty
Grupa 13 protofilamentów łączy się wzdłuż długiej osi tworząc ścianę jednej
mikrotubuli
Polimeryzacja i depolimeryzacja mikrotubul jest spontaniczna i zachodzi bardzo
szybko

48
Do przeprowadzenia polimeraz potrzebne jest GTP i jony Ca
2+
Hydroliza GTP jest potrzebna do odkształcenia cząsteczki mikrotubuli, a nie od
przeprowadzenia polimeryzacji
Ruch mikrotubul, a wraz z nim przemieszczanie się organelli, powoduje
polimeryzacja, która zaczyna się na biegunie „+” i depolimeryzacja na biegunie „-„
(zachodzą one jednocześnie)
Polimeryzacja rozpoczyna się w pobliżu centrioli w cytoplazmie
Niektóre alkaloidy, takie jak
kolchicyna,
hamują polimeryzację mikrotubul, co
przyczynia się do zatrzymania mitozy z metafazie
Związki te to są tzw. antymitotyki
Mikrotubule występują we wszystkich komórkach, tworzą rzęski (9+2), witki,
wrzeciono podziałowe i centriole
Poza tym mikrotubule występują jako proste, niezaginające się struktury decydujące o
kształcie komórki
Mikrotubule mogą wiązać się z białkami MAP
Białka te wpływają na polimeryzację i depolimeryzację mikrotubul, i należą do nich:
MAP – 2 i białko tan (z grupy MAP), które zapobiegają depolimeryzacji,
zapewniając stabilizację równoległych mikrotubul w dendrytach i aksonach.
Nadmierna depolimeryzacja białek tan doprowadza do bezładnego ułożenia
mikrotubul, uniemożliwiając transport wzdłuż aksonu
Białka motorowe (z grupy MAP), takie jak inozyna, dyneina. Cząsteczki tych
białek mają podobną budowę do miozyny. Jednym końcem wiążą się z
transportowaną strukturą, a drugim oddziaływają z mikrotubulą. Mają
zdolność ATP-azy. Pod wpływem ATP mogą zmieniać kąt ułożenia cząsteczek
względem mikrotubuli wywołując ruch rzęsek i witek
Miejsce powstawania mikrotubul:
Brak jest wyraźnych organizatorów w komórkach roślin wyższych (brak
centrioli, ciałek podstawowych), jednakże podczas cyklu życiowego komórek
roślinnych, mikrotubule polimeryzują w stałych miejscach, tzw. centrach
nukelacji mikrotubul (MTOC)
W interfazie cyklu życiowego komórek, MTOC można wykrywać metodami
immunocytochemicznymi i stwierdzono, że występują na powierzchni jądra
komórkowego, na błonie, plastydach
Największą rolę w polimeryzacji mikrotubul pełni otoczka jądrowa –
mikrotubule tu wytwarzane rosną do cytoplazmy, a nawet do błony
komórkowej
Centrosfera i centriole
Występują u zwierząt
Są to struktury cytoplazmatyczne o kształcie walcowatym
Każda
centrofera
składa się z centrioli i otaczającej ją cytoplazmy
W cytoplazmie dookoła centrioli znajdują się kompleksy tubuliny γ i innych białek,
które są ośrodkami polimeryzacji mikrotubul
Każda centriola zbudowana jest z 9 trójek mikrotubul ułożonych koncentrycznie
W środku znajduje się filament i niewielkie ilości DNA
Poszczególne trójki mikrotubul łączą się ze sobą za pomocą
białka fibrylarnego
W komórkach niedzielących się, znajdują się dwie centriole, których długie osie są w
stosunku do siebie pod kątem prostym
Przed podziałem komórki, w fazie S, zachodzi replikacja centrioli i ich DNA

49
Powstają w ten sposób dwie pary centrioli, które wędrują do dwóch różnych biegunów
komórki
Centrosfera i znajdujące się w niej centriole, biorą udział w organizacji biegunowej
struktury wrzeciona kariokinetycznego oraz biorą udział w polimeryzacji mikrotubul
tego wrzeciona
Występują też jako kinetosom, biorący udział w strukturze mikrotubularnej i
synchronizowaniu ruchu rzęsek
Filamenty pośrednie:
Mają średnice około 10 nanometrów (8-11 nanometrów)
Są rozmieszczone w całej cytoplazmie komórki
Występuja pojedynczo lub tworzą siei, albo pęczki
Filamenty są bardzo stabilne i wytrzymałe na rozciąganie i bardzo odporne na
działanie związków chemicznych
Nadają one komórkom i tkankom dużą odporność mechaniczną
Są splecionymi włóknistymi polipeptydami, składającymi się z polipeptydów
fibrylarnych połączonych ze sobą powierzchniami bocznymi
Wielkość tych polipeptydów jest różna
Ich masa cząsteczkowa wynosi około 40 – 200000 i zależy od rodzaju filamentu i
rodzaju komórki
Na ogół jedna komórka ma jeden typ filamentów pośrednich
Mamy 6 typów filamentów pośrednich:
1. filamenty cytokeratynowe kwaśne
inaczej tonofilamenty/tonofibryle
2. filamenty cytokeratynowe zasadowe
ze względu na dużą ilość białek je budujących, są najbardziej różnorodną klasą
pośród filamentów pośrednich
występują głównie w komórkach nabłonkowych, szczególnie obficie w
narażonych na działanie mechaniczne (komórki naskórka)
3. filamenty wimentynowe, desminowe, glejowe
występują w komórkach tkanki łącznej
4. filamenty neurokeratynowe (neurofilamenty)
występują w większości w komórkach nerwowych
5. filamenty laminowe blaszki jądrowej
laminy jądrowe tworzą sieć gęstych filamentów pośrednich znajdujących się
tuż pod osłonką jądrową
6. filamenty festynowe w rozwijających się neuronach
budują filamenty pośrednie w neuronach zarodków i w mięśniach
o
Są charakterystyczne dla poszczególnej tkanki
o
Wykorzystywane są w badaniach immunocytochemicznych przez histologów do
rozpoznawania rodzajów tkanek
17.01.2008
Regulacje cyklu komórkowego
G0 – komórki mogą wyjść z fazy G1, odbywa się różnicowanie
cykl komórkowy – cykl mitotyczny lub cykl podziałowy – szereg kolejno następujących po
sobie zmian chemicznych, fizycznych i strukturalnych komórki prowadzący do podwojenia

50
materiału genetycznego i precyzyjnego rozdziału między dwie potomne komórki. Składa się z
interfazy: G1, S, G2 oraz fazy H
Synteza DNA – podwajanie jądrowego DNA w fazie S cyklu trwa 6-8 godzin. Reperacja
(=synteza uszkodzonego fragmentu) odbywa się w całej interfazie. Nie zachodzi ponowna
replikacja DNA w tej samej fazie S. Synteza mitochondrianlnego DNA odbywa się także
poza fazą S.
Mitoza (faza M) = kariokineza + cytokineza
- Kariokineza (P, M, A, T) Kondensacja chromatyny -> rozejście się siostrzanych
chromatyd na bieguny komórek, wytworzenie i rozpad wrzeciona podziałowego,
fragmentacja oraz odbudowa otoczki jądrowej i jąderka.
- Wrzeciono podziałowe – dwubiegunowa struktura zbudowana z mikrotubuli
kinetochorowych, ramiennych, biegunowych i astralnych oraz białek. Wrzeciono
tworzą centrosomy z centromerami oraz specjalne białko mikrotubuli i cytoplazmy.
Odpowiada za ułożenie chromosomów w płaszczyżnie równikowej komórki oraz za
odprowadzanie ku centromerom. Wpływa też na wielkość komórek potomnych oraz
ich zróżnicowanie.
Autonomiczna regulacja cyklu komórkowego – ma charakter kaskadowy
i jest regulowany
wieloma genami wytwarzającymi określone białka. Poszczególne fazy są ułożone w stałej
kolejności – następna faza rozpoczyna się po pomyślnym zakończeniu poprzedniej:
- geny kodujące białka pobudzające cykl to protonkogeny
- geny kodujące białka hamujące cykl to geny supresorowe.
Kliniczną rolę odgrywają kinazy zależne od cyklin CDK (8), które są aktywowane przyz
cykliny (14) kwasy
zależne od cyklu
Stężenie CDK w komórkach jest stałe, cykliny zmieniają
się w różnych fazach, co powoduje
zmianę aktywności różnych CDK w czasie cyklu -> zakończenie jednej fazy przyczyniło się
do zakończenia drugiej.
Obok CDK istnieją również inhibitory cyklu CKI = hamujące cykl komórkowy.
Należą do
rodziny białek p21, INK4, RB i p53
Punkty kontrolne cyklu komórkowego – regulują przejścia do następnej fazy. Są to:
1. punkt kontrolny późnej fazy G1 (punkt restrykcyjny) – decyduje o wejściu komórki do
cyklu komórkowego (interfaza)
2. punkt kontrolny późnej fazy G2 – decyduje o wejściu komórki w mitozę (po usunięciu
detektorów DNA) (inetrfaza)
3. punkt kontrolny wrzeciona podziałowego – decyduje o związaniu kinetochorów
wszystkich chromosomów z mikrotubulami wrzeciona, zapobiega powstaniu aneuploidów
(anafaza)
Regulacja cyklu komórkowego z zewnątrz:
- Czynniki zewnętrzne działają zarówno na komórkę w cyklu komórkowym i w fazie G0 (w
stanie spoczynku)
- Wpływają na rozpoczęcie i czas trwania cyklu

51
Są to cytokininy, które wiążą się z receptorami błonowymi ->
uaktywowanie
białek
RAS -> uaktywnienie kinazy
MAPERK -> fosforylacja białek -> synteza nukleotydów
-> ekspresja protoonkogenów
-> wejście komórki do cyklu.
Choroby proliferacyjne:
- choroby nowotworowe (nadmierna ekspresja protoonkogenów)
- choroby sensowo-naczyniowe (miażdżyca, nadmierna proliferacja miocytów naczyń
krwionośnych
- choroby wirusowe
Opis cyklu komórkowego:
czas trwania:
8 min komórki larw owadów
30 min – u bakterii
4 – 12 godz – wczesne stadia rozwoju ssaków
24 godz – większość komórek ssaków
ponad rok – komórki trzustki
- Faza G1 – trwa krótko- kilkanaście godzin = synteza DNA, białek strukturalnych,
regulatorowych i enzymatycznych.
Pod koniec fazy G1 – punkt restrykcyjny (kontrolny)
czyli zgromadzenie zestawu odpowiednich białek regulatorowych i enzymatycznych
potrzebnych do rozpoczęcia fazy S.
unieczynnienie białek hamujących przejście przez ten punkt.
Niekiedy z fazy G1 komórka przechodzi z cyklu komórkowego w stan spoczynku - G0.
- Faza S – synteza DNA rozpoczyna się w punktach startowych replikacji DNA (w kilkunastu
tożsamych miejscach)
euchromatyna wcześniej, a heterochromatyna później
dwukierunkowo z prędkością 10-100 nukleotydów/s (1,8 m DNA jest zreplikowane w
ciągu 6-8 godzin)
helikaza rozczepia podwójną nić DNA - powstają widełki replikacyjne
polimeraza –> synteza DNA
Replikacja DNA w tym samym cyklu komórkowym zachodzi tylko 1 raz.
- Faza G2 – ok. 2–4 godz.
Synteza RNA, białek regulatorowych i enzymatycznych potrzebnych do wejścia
komórki w mitozę
Dodatkowo jest syntetyzowana nowa błona komórkowa potrzebna do zużywania w
czasie cytokinezy(λ
nowe komórki potomne mają większą
powierzchnię).
Komórki wychodzące z tej fazy muszą sforsować punkt kontrolny G2 – uszkodzenia genomu
muszą być naprawione.
Mechanizmy naprawy:
1. rozpoznanie uszkodzonego fragmentu DNA i wycięcie przez nukleazy
2. uzupełnienie tego fragmentu przez polimerazę DNA (na podstawie komplementarnej
nieuszkodzonej nici)
3. wbudowanie przez ligazę nowego fragmentu do naprawionej nici.
52
Mitoza (=faza M) u ssaków trwa 30-180 min.
Kariokineza:
- Profaza – kondensacja chromatyny w wyniku fosforylacji histonu H1 przez kinazy fazy M
oraz defosforylacji histonów H3.
Powstanie wrzeciona podziałowego (mikrotubule kinetochorowe, ramienne,
biegunowe, astralne)
Za utrzymanie kształtu wrzeciona oraz ruchy chromosomów odpowiadają białka
motorowe: kinezyna i dyneina cytoplazmatyczna
Wytworzenie wrzeciona podziałowego poprzedza podział centrioli i centrosomów
Fragmentacja otoczki jądrowej i jąderka (zanik obu) (Udział kinazy białkowej
fosforylującej białka otoczki jądrowej i jąderka)
-Metafaza – chromosomy ustawiają się w płaszczyźnie równikowej komórki, chromatydy są
położone w 1 punkcie w centromerach -> przemieszczanie składników cytoplazmy do
biegunów
-Anafaza – rozdział chromatyd i ich rozsunięcie do biegunów komórki w skutek
uaktywniania sekuryny przez kompleks anafazowy APC. Ruch chromatyd następuje za
pomocą kilku mechanizmów:
Ślizgania się względem siebie mikrotubul biegunowych (udział kinezyn), mikrotubul
astralnych (udział dyneiny), mikrotubul ramiennych (udział chromokinezyn),
kinetochorów (kinezyny i dyneina cytoplazmatyczna)
Depolimeryzacja mikrotubul kinetochorowych na końcach komórki
- Telofaza – dekondensacja chromosomów w wyniku defosforylacji histonu H1
Rozpoczęcie transkrypcji DNA -> synteza mRNA
Odtworzenie jąderka i otoczki jądrowej wskutek defosforylacji lamin
Zanik wrzeciona podziałowego (polimeryzacja mikrotubul przez tubulinę)
Cytokineza - podział cytoplazmy wskutek wytworzenia pierścienia kurczliwego z filamentów
aktynowych i miozynowych i jego skurczu
............
GTP-azy oraz fosfataza łańcuchów
lekkich miozyn
Mechanizm skurczu:
1. z udziałem Ca2+
2. depolimeryzacja i polimeryzacja filamentów
Mitoza asymetryczna – powstałe 2 komórki potomne różnią się między sobą – jedna jest
identyczna jak matka, a druga różnicuje się.
Mechanizm może podlegać:
niesymetrycznej lokalizacji białek NUMB i PROSPERO (występują tylko na jednym
biegunie)
niesymetrycznie rozmieszczenie mRNA dla tych białek
Sposoby badania cyklu:
1. Metoda statmokinezy ( hamowanie mitoz w metafazie np. przy pomocy kolchicyny) –
określanie odsetka komórek mitotycznych w stosunku do interfazowych
2. Metoda autoradiografii – podawanie znakowanych nukleotydów, które w fazie S są
wbudowywane do jąder

53
3. Cytometria przepływowa – znakowanie DNA jądrowego fluorochromem -> rozkład
DNA w poszczególnych komórkach – określenie czasu trwania fazy G1+S; G2-M
Regulacja cyklu komórkowego:
1. Znaczenie kinazy białkowej.
Prowadzony cykl komórkowy jest regulowany przez fosforylację białek enzymatycznych,
które je uaktywnia. Fosforylację przeprowadzają enzymy CDK – kinazy zależne od cyklin
(fosforyluje grupy serynowe i treoninowe białek)
W różnych fazach cyklu są różne CDK i są aktywowane przez rodzine białek nazywanych
cyklinami.
-stężenie CDK jest stałe, a stezenie cyklin zmienia się cyklicznie
-aktywacja CDK polega na tworzeniu kompleksu z odpowiednią cykliną
-dotychczas poznano 8 rodzajów CDK (CDK- 1-8) .
...
cyklu oznaczamy literami A-J
2. Naturalne inhibitory cyklu.
W komórkach istnieja specjalne punkty kontrolne cyklu, w których przechodzenie przez
cykl może być hamowane na pewien czas lub trwale ( wejście w stan spoczynkowy G0).
Hamowanie cyklu umożliwia usunięcie zaburzeń lub uśmiercenie komórki wadliwej przez
apoptozę.
Są to inhibitory CDK, CDI – uniemożliwiają wiązanie się cyklin z kinazami lub blokują
wiązanie kompleksu cyklin (CDK z substratami)
Istnieją 4 rodziny CDI:
Białko 21 (+p27 i p57)
Rodzina białek INK4
(+ białka p15, p16, p17, p19)
Rodzina białek RB – białka te są produktem genu rb, który zatrzymuje
komórki w fazie G1, hamuje transkrypcję odpowiednich genów, których białka
są potrzebne do przejścia z G1 do S. Jest nazywane wrotami cyklu
komórkowego -> jego fosforylacja (inaktywacja pozwala na wejście do cyklu)
Rodzina białek p53 – białko p53jest czynnikiem transkrypcyji, hamuje
komórki w fazie G1, pozwala na naprawę DNA bądź na apoptozę. Nazywane
jest strażnikiem genomu.
Cykl autonomiczny – występuje we wczesnych stadiach rozwoju zarodkowego, trwa od 8
min do 12 godzin
-cykliczne zmiany stężenia cyklin
-brak czynników zewnętrznych
-nie ma wzrostu komórki, fazy: G1 i G2 są krótkie lub nie ma ich w ogóle
Przejście G1 -> S regulują kinazy CDK4 i CDK6, które są aktywowane przez cykliny D1,
D2, D3.
Przejście przez punkt restrykcyjny fazy G1 umożliwia aktywność kompleksu cykliny
CDK przy niskim stężeniu CKI – p27 oraz nieczynnym białku RB
Czynniki fazy S (SPF) i kompleks cyklina E/CDK2 i cyklina A/CDK2, umożliwiają
rozpoczęcie i zakończenie syntezy DNA i wejście do kolejnego etapu interfazy G2.
Przejście G2 -> M
Rozpoczęcie mitozy zależy od kompleksu cyklin B1, B2/CDK1, które są nazywane MPF. Do
tego jest potrzebna podwójna fosforylacja i defosforylacja CDK1, która w kompleksie z

54
cyklinami B1 i B2 są w pełni aktywne i fosforylują wiele białek
jądrowych – kondensacja
chromatyny, zanik jąderka i otoczki jądrowej.
Wyjście komórki z mitozy – regulowane jest przez zestaw białek wyjścia z mitozy – MEN
powodujących inaktywację kompleksu cyklina B/CDK1. Białka te zapobiegają wchodzeniu
do fazy G1 komórek z defektami.
Czynniki zewnętrzne
Mogą zmieniać czas cyklu lub zmuszać koórkę do wejścia do cyklu. Są to peptydowe
czynniki wzrostu różnicowania, czyli cytokiny. Jest ich ponad 1000, najlepiej poznano PDGF,
FGF, TGF, NGE, TNF oraz ponad 20 interlenkin. Cytokiny łączą się z białkowymi kinazami
MAPERK.
Kinazy MAPERK:
-pobudzają syntezę
nukleotydów (do syntezy DNA)
-pobudzają transkrypcję protoonkogenów
-ułatwiają tworzenie się kompleksów cyklin D1 (CDK4, który unieczynnia
białko RB) ->
aktywacja transkrypcji kilku protoonkogenów.
-fosforylacja histonu 3, który rozluźnia chromatynę i umożliwia transkrypcję.
Cytokiny zmuszają komórkę z fazy G0 do wejścia w cykl komórkowy.
Wpływ podłoża i adhezji komórek
Adhezja komórek do podłoża wielu komórek jest niezbędnym warunkiem do rozpoczęcia i
kontynuacji cyklu komórkowego - > hamuje aktywność inhibitorów CKI, a szczególnie
białko p21, p27 oraz unieczynnia białko RB (hamuje cykl komórkowy).
Choroby proliferacyjne:
1.Choroby nowotworowe – wskutek niekontrolowanej proliferacji.
Defekt polega na łatwym przechodzeniu przez punt restrykcyjny fazy G1. 50% przypadków
to skutek nadmiaru cykliny D1, która może pobudzać transformację nowotworową nabłonka
sutka
2.Choroby sercowo-naczyniowe – nadmierna proliferacja komórek mięśni gładkich –
powstanie w nich guzków oraz ich degeneracja i wapnienie. Jest to wpływ cytokin PDGF.
Choroby wirusowe- niektóre nowotwory współistnieją z zakażeniami wirusowymi komórek,
np. wirusem ludzkich
brodawczaków (HPF). Wirusy hamują CKI – białko p21 i p27,
prowadzi również do rozkładu białko p53 oraz niskie położenie między białkiem RB, a
czynnikiem transkrypcji E2F, co pobudza cykl komórkowy. Dodatkowo wirusy uczynniają
geny kodując cykliny A i E pobudzając cykl komórkowy.
Wyszukiwarka
Podobne podstrony:
biologia nmr id 87949 Nieznany
biologia pp o id 88225 Nieznany (2)
czynniki biologiczne 10 id 6672 Nieznany
biologia wyklady id 88204 Nieznany (2)
biologia ABCDE id 87704 Nieznany
Biologia Wyklad 9 id 88128 Nieznany (2)
Biologia nr 3(1) id 87951 Nieznany
Biologia Cwiczenia 4 id 87712 Nieznany (2)
Biologia Cwiczenia 8 id 87713 Nieznany (2)
biologia 2008 id 87683 Nieznany (2)
Biologia Cwiczenia 9 id 87714 Nieznany (2)
BUDOWA KOMoRKI id 94229 Nieznany (2)
biologia spektroskopia2 id 8805 Nieznany
biologia mol id 87896 Nieznany
Biologia Wyklad 6 id 88126 Nieznany (2)
biologia molekularna id 87900 Nieznany (2)
biologia synteza id 88080 Nieznany
Biologia 10 id 87672 Nieznany
więcej podobnych podstron