„Rozwój układu nerwowego”
(WBNZ – 818)
Jolanta Górska-Andrzejak
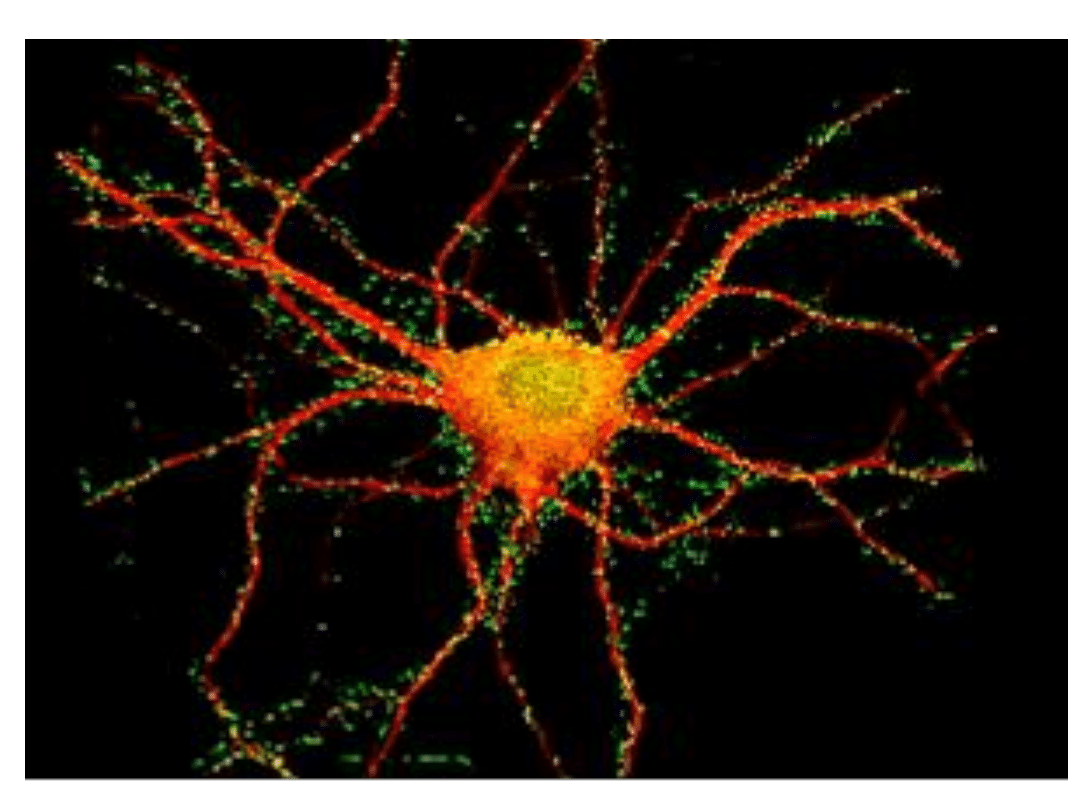
Neuron hipokampa szczura w hodowli komórkowej. Czerwony –tubulina, Zielony- synapsyna.
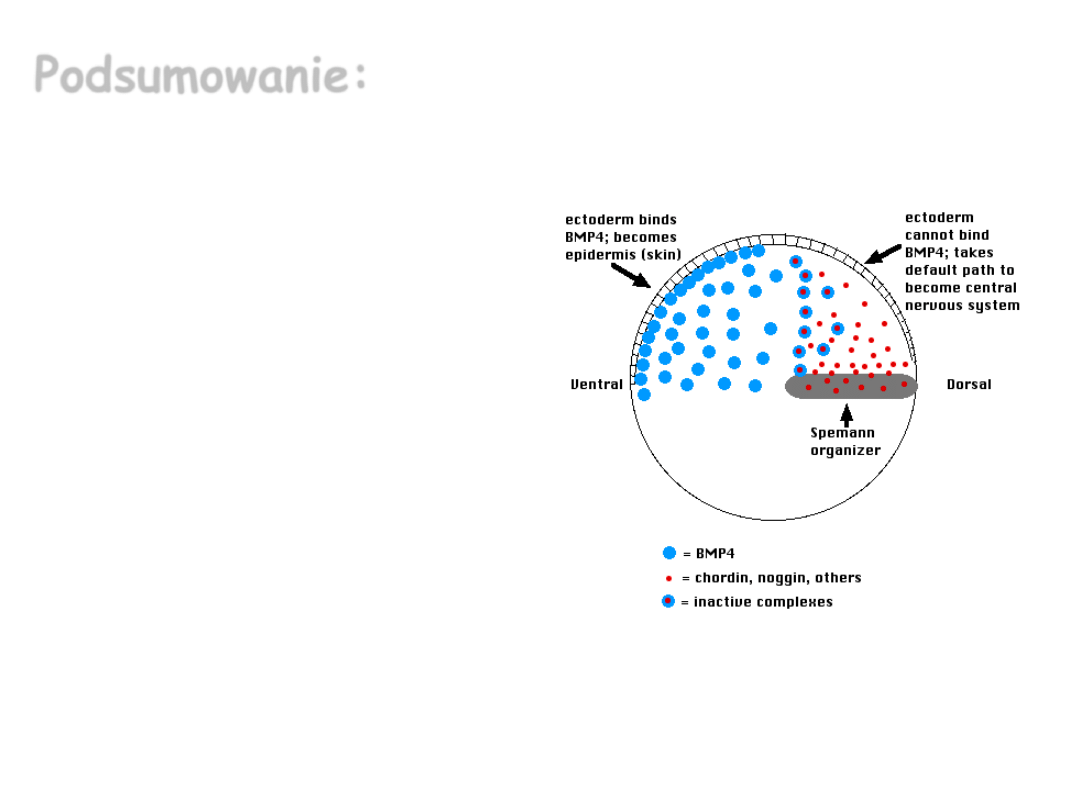
Podsumowanie:
Badania z zakresu embriologii
klasycznej wykazały, że układ
nerwowy wykształca się w
czasie rozwoju embrionalnego
z ektodermy na skutek
indukcji ze strony tkanki
budującej tzw. organizator
Spemann’a. Dalsze badania
pozwoliły poznać także
molekularny mechanizm
indukcji układu nerwowego,
polegający na inhibicji ścieżki
sygnałowej BMP (zależnej od
BMP represji) przez induktory
neuronalne.
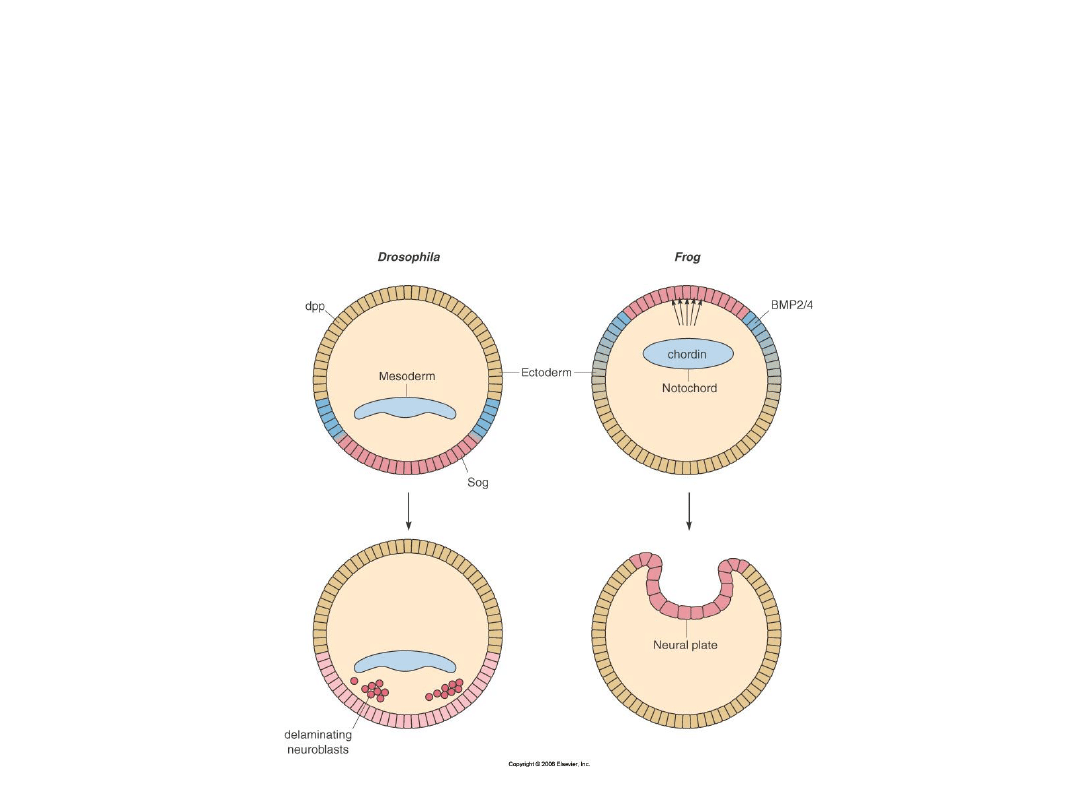
dpp
zapobiega
różnicowaniu
w neuroektodermę
Sog
promuje
różnicowanie
w neuroektodermę
dpp, Sog
BMP2/4,Chordina
BMP
zapobiega
różnicowaniu
w neuroektodermę
Chordina
promuje
różnicowanie
w neuroektodermę
• Zarówno u
Drosophila
jak i u
Xenopus
do wykształcenia się w
embrionie regionu neurogenicznego konieczny jest antagonizm
czynników TGF-beta, takich jak dpp/BMP, przez substancje będące
induktorami neuronalnymi, które są wytwarzane przez organizator.

Wytwarzane przez organizator
noggina, chordina czy follistatyna
zapobiegają łączeniu się BMP do ektodermy i mezodermy w
pobliżu organizatora – > zapobiegają indukcji/powstaniu
w tym rejonie epidermy - zezwalają na powstanie tkanki
nerwowej.
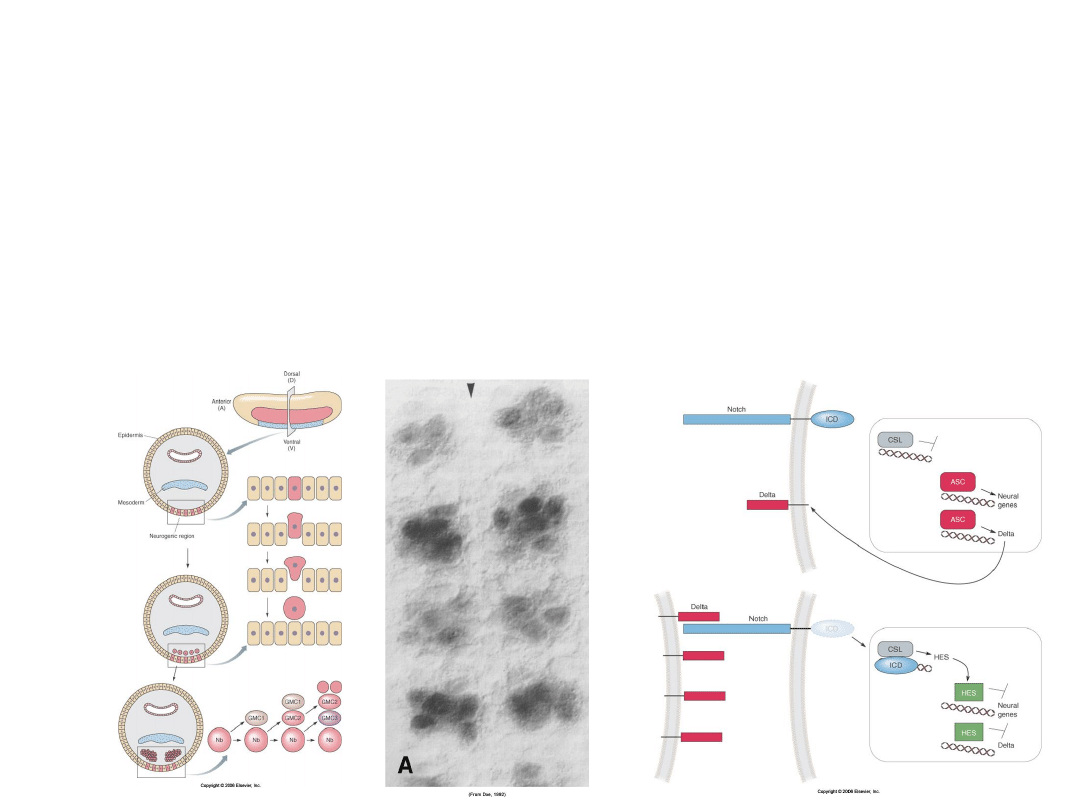
Antagonizm BMP jest konieczny i wystarczający do indukcji
układu nerwowego, jednakże u
Drosophila
proces delaminacji
neuroblastów z regionu neurogenicznego (neuroektodermy)
zachodzi dzięki drugiej ścieżce sygnałowej –>
Notch/Delta,
która reguluje ekspresję i funkcję białek proneuralnych
będących czynnikami transkrypcji
bHLH
.

Proneuronalny system
Notch/Delta jest konserwatywny i
występuje również u kręgowców

Wykład 3
Segmentacja układu nerwowego
Różnicowanie zależne od pozycji
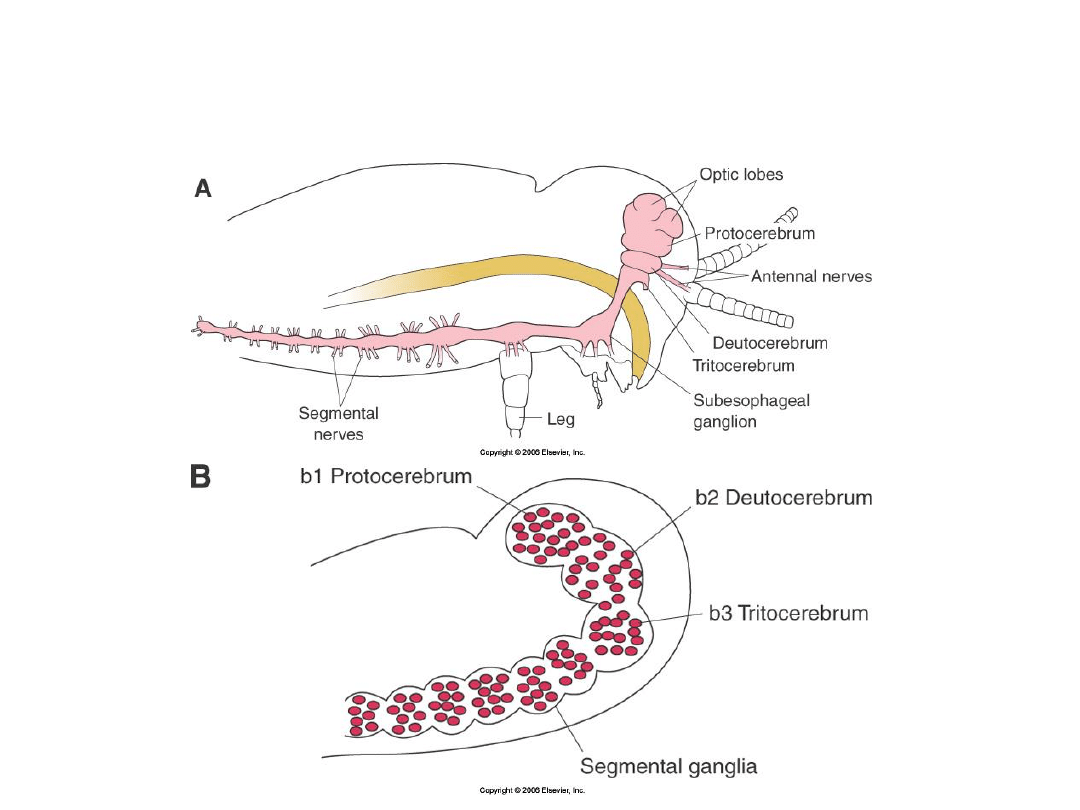
Układ nerwowy
D. melanogaster
Różnicowanie zależne od pozycji

Model Flagi Francuskiej
(Wolpert, 1969, 1978)

Kształtowaniem się zróżnicowanej budowy ciała
wzdłuż osi przednio-tylnej
u
Drosophila
zawiaduje hierarchiczny układ
genów regulatorowych.

Plan budowy ciała zostaje zdeterminowany już w
stadium blastodermy
-> uaktywnienie genów wyznaczających pozycję i
organizację przyszłych segmentów ciała.
• 5 grup genów tworzy hierarchiczny
układ regulacyjny.
• Geny wyższej kategorii decydują o
aktywności genów podrzędnych.
• Geny w obrębie tej samej kategorii
mogą nawzajem wpływać na swoją
aktywność
.

geny polarności jaja
(decydują o orientacji przedniej i tylnej części ciała;
pozycja nadrzędna)
geny segmentacji
(3 kategorie; wyznaczają podział ciała na segmenty)
geny homeotyczne
(decydują o prawidłowym zróżnicowaniu budowy każdego
segmentu zgodnie z jego pozycją wzdłuż osi przednio-
tylnej)
Geny te kodują białka typu regulatorowego, których struktura
wskazuje na to, że mogą wiązać się ze specyficznymi
sekwencjami DNA i jako czynniki transkrypcji wpływać na
aktywność innych i własnych genów.

Geny polarności jaja
(ang. egg polarity genes)
• Wywołują tzw.
efekt mateczny
,
gdyż ich
produkty zostają zdeponowane w oocycie,
są więc pod kontrola genotypu matki
(pod kontrolą genów matczynych; ang. maternal genes)
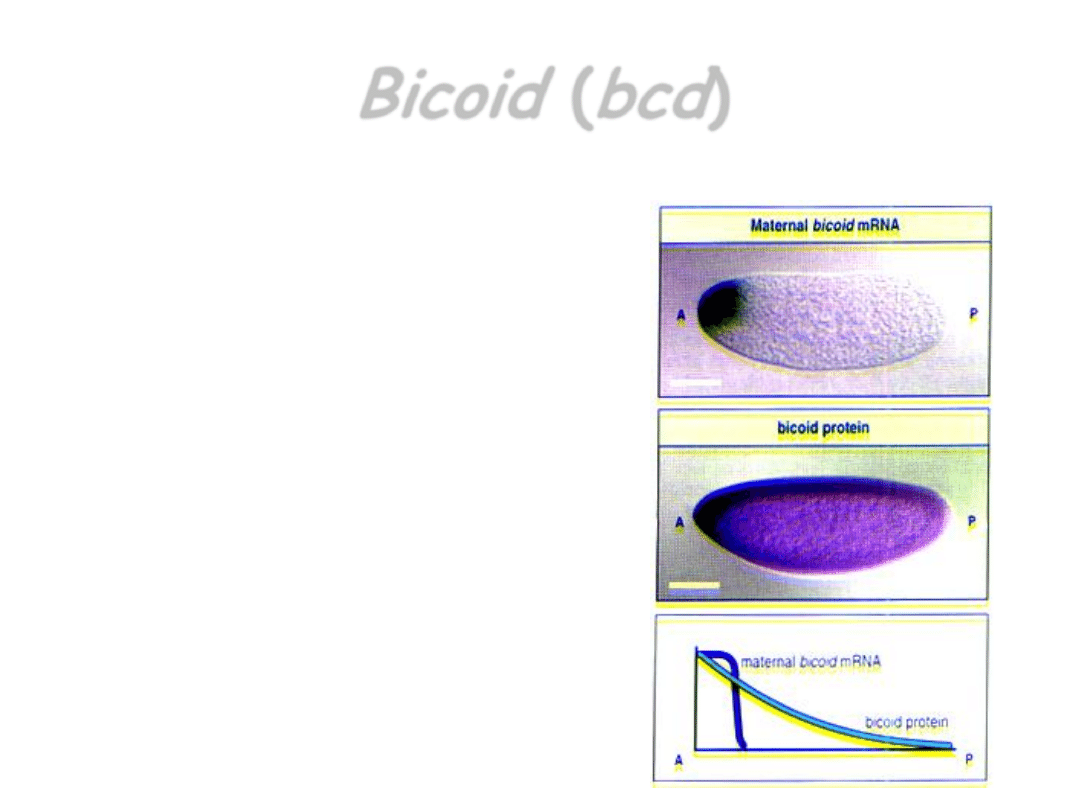
Bicoid
(
bcd
)
wyznacza
przedni
koniec ciała
• cząsteczki mRNA genu
bicoid
są syntetyzowane
w organizmie matki, w
komórkach odżywczych
jajnika, skąd przechodzą
do oocytu i gdzie od razu
zostają zakotwiczone
• po zapłodnieniu jaja na
matrycach tego mRNA
syntetyzowane jest
białko, które
rozprzestrzenia się i
tworzy gradient stężenia,
malejący ku tyłowi.
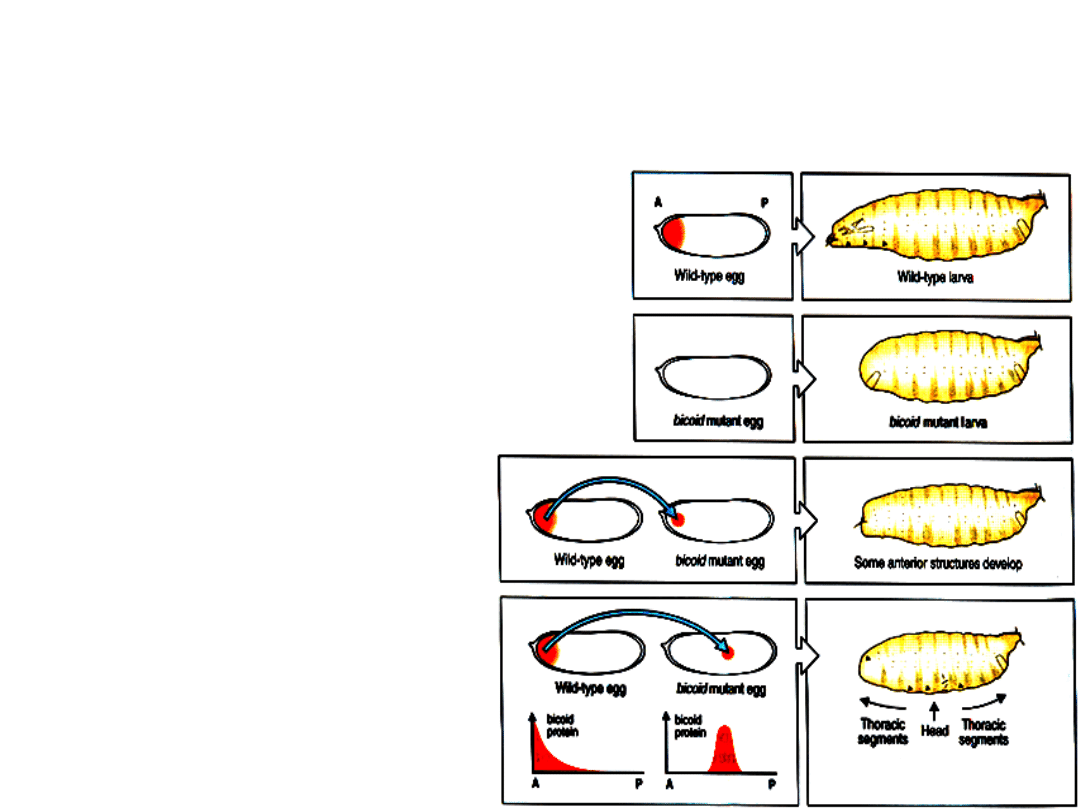
Zarodki mutantów
bcd
pozbawione są
głowy i struktur tułowiowych
bicoid
zachowuje się jak
typowy morfogen, (tzn.
substancja wywołująca
specyficzną indukcję);
cytoplazma zawierająca
białko kodowane przez ten
gen po wprowadzeniu w
dowolną okolicę mutanta
bicoid
indukuje w tym
miejscu powstanie struktur
charakterystycznych dla
przedniej części ciała.

Nanos (nos)
gen wyznaczający tylny koniec jaja, niezbędny do powstania
segmentów
odwłokowych.
cząsteczki mRNA tego genu są
syntetyzowane w organizmie
matki.
ulega translacji w tym samym
czasie co
bicoid
.

• Syncytialna blastoderma zawiera przynajmniej
dwa przeciwstawne gradienty produktów
kodowanych przez geny polarności jaja, takich
jak
bicoid
i
nanos
.
• Białka Bicoid i Nanos dostarczają wstępnych
sygnałów pozycyjnych, które wpływają na
aktywność genów niższej kategorii (genów
segmentacji) zawężając ich ekspresję do
konkretnych regionów ciała.

hunchback i caudal
W jaju występują także matczyne transkrypty genów
hunchback
i
caudal
.
•
hunchback
,
tak jak
bicoid
wyznacza przedni
koniec ciała
•
caudal
, tak jak
nanos
, wyznacza tylny koniec
ciała.
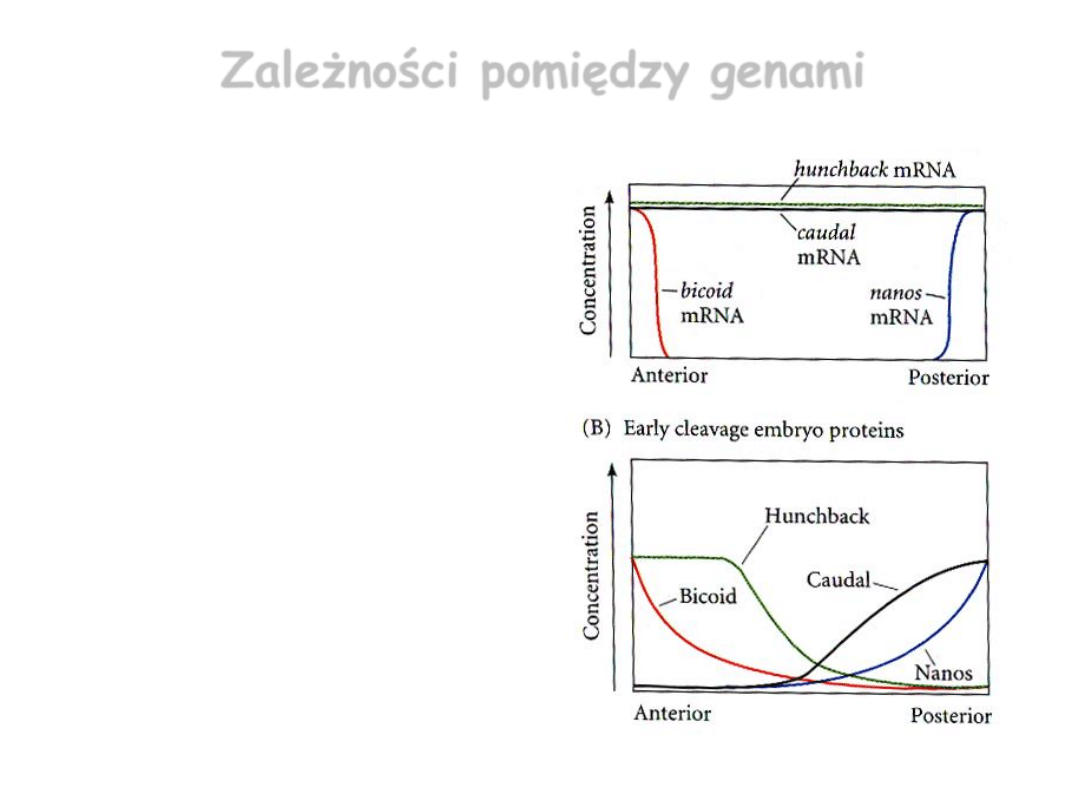
Zależności pomiędzy genami
• Bicoid zapobiega translacji
caudal
w przedniej części jaja.
• Nanos powoduje inhibicję
translacji
hunchback
w tylnej
części jaja.
• Gradient
hunchback
jest
dodatkowo wzmocniony dzięki
zachodzącej transkrypcji tego
genu w przedniej części jaja=>
białko Bicoid jest czynnikiem
transkrypcji
hunchback
.

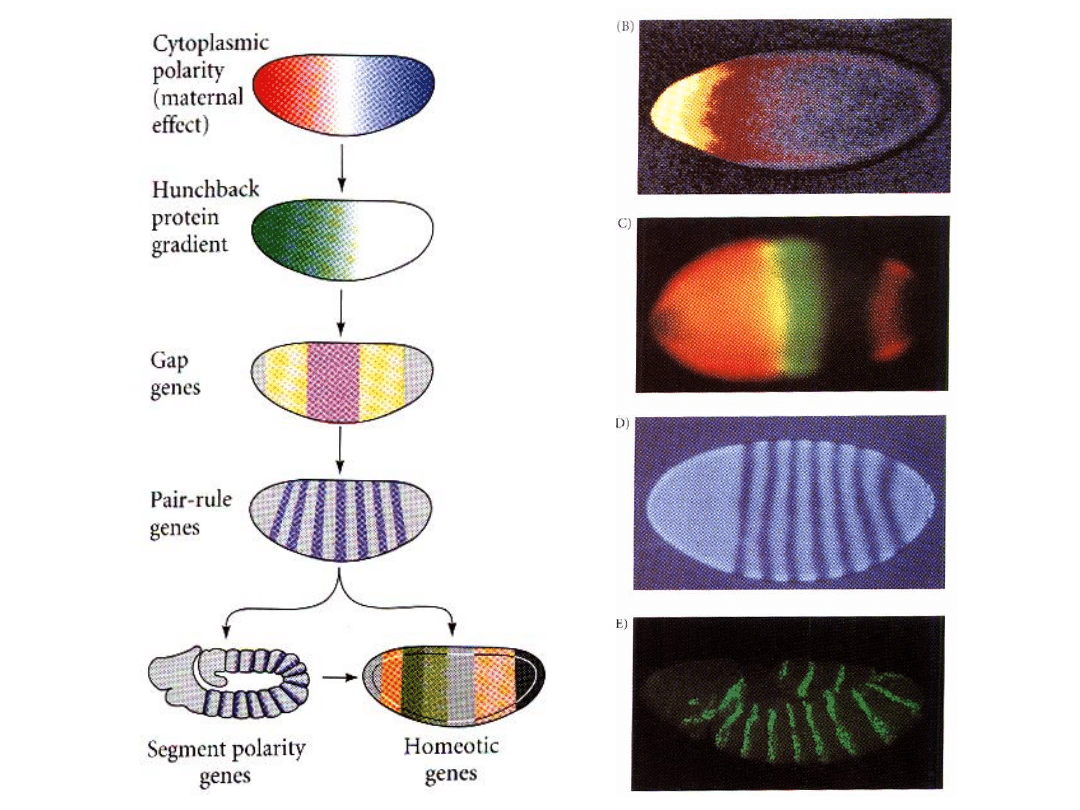
Geny Luki
(ang. GAP genes)
-pierwsze geny zygotyczne, które ulegają ekspreji wzdłuż osi
przednio-tylnej
Geny segmentacji, których mutacje
wywołują utratę dużych partii ciała (ang.
gap genes) i wczesną letalność embrionu.
(pierwsza kategoria genów segmentacji)
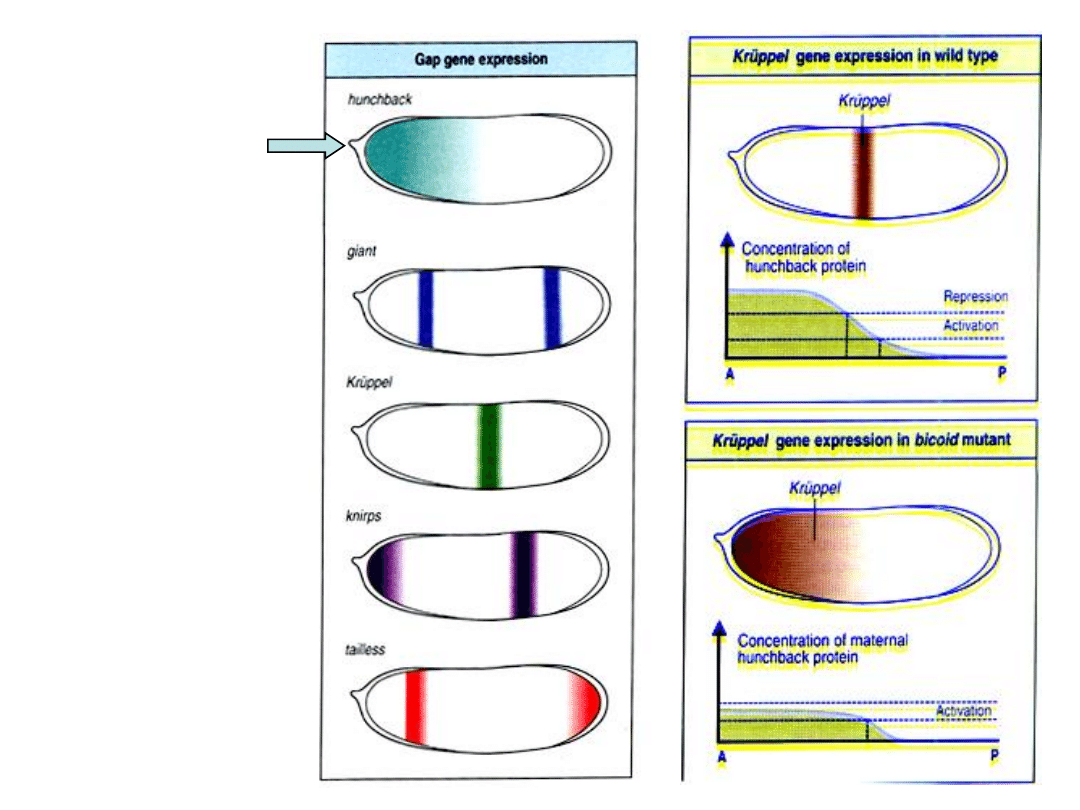
Np.
Kr
ǜ
ppel
–
ekspresja tego genu
zachodzi tylko w
środkowej części
embrionu
(zarówno w części
przedniej jak i w
tylnej jego
transkrypcja jest
hamowana przez
białka genów
polarności jaja).
Białka Bicoid,
Hunchback i Caudal
są czynnkami
transkrypcyjnymi, które
aktywują geny luki

Geny reguły
parzystej
(geny parzystości segmentów,
ang.pair-rule genes)
• geny segmentacji o bardziej lokalnym działaniu, wyznaczające
regiony tzw. parasegmentów
(zaczątków segmentacji, które
są przesunięte w stosunku do ostatecznych segmentów; 1/3
tylnej części segmentu + 2/3 segmentu następnego )
• ich mutacje wywołują brak co drugiego parasegmentu, np.
mutanty
fushi tarazu
(po japońsku: mało segmentów,
ftz
) nie
mają parasegmentów nieparzystych (brązowy), natomiast
mutanty
even-skipped
(
eve
) parzystych (fioletowy).
• Ich ekspresja zbiega się w czasie z celularyzacją;
przekształcenie blastodermy syncytialnej w komórkową.
(druga kategoria genów segmentacji)

Geny polarności segmentów
(ang. segment polarity genes)
(trzecia kategoria genów segmentacji)
Wyznaczają określoną część każdego
segmentu. Ich mRNA wykrywane jest w
formie prążków, często o szerokości jednej
komórki.
(trzecia kategoria genów segmentacji)
Ulegają ekspresji w blastodermie komórkowej, w postaci 14 prążków
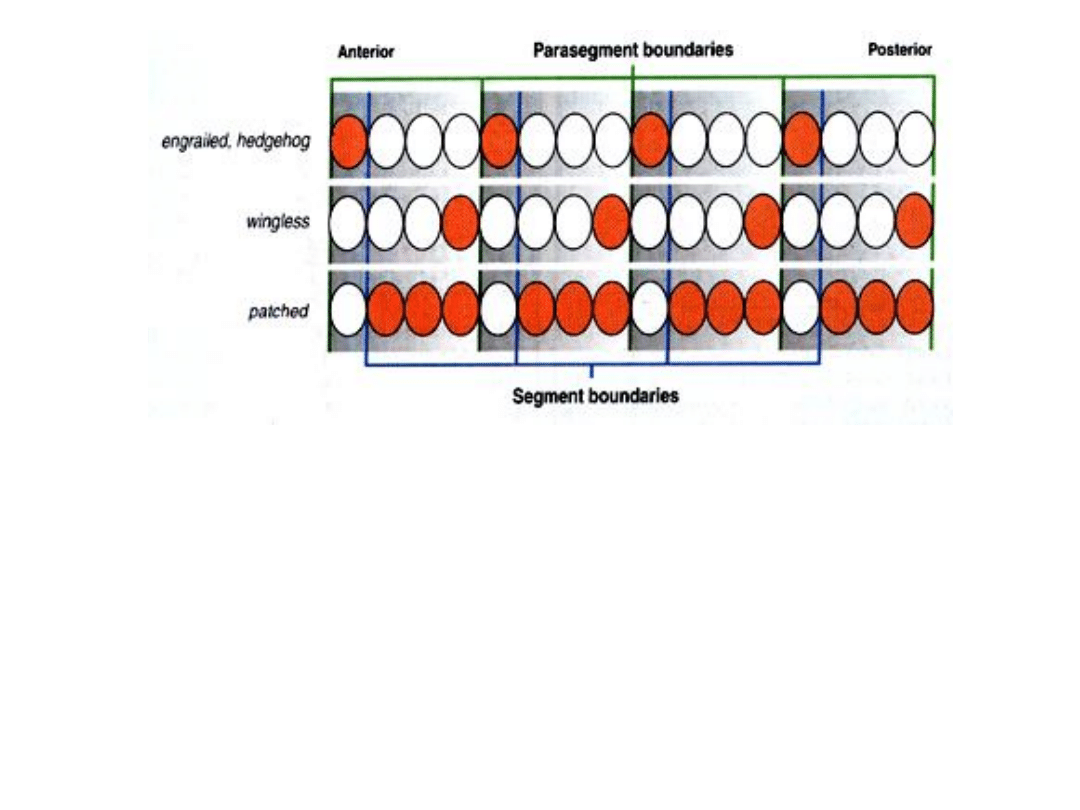
• mRNA genu
engrailed
(
en
), wykrywalny jest w formie 14 prążków,
każdy o szerokości jednej komórki, zlokalizowanych w przedniej
ćwiartce poszczególnych parasegmentów.
• mRNA genu
wingless
(
wg
) – w ćwiartce tylnej. U mutantów, ta część
każdego parasegmentu zbudowana jest jak lustrzane odbicie części
pozostałej.
• Po gastrulacji gen
patched
ulega ekspresji, ale tylko w tych
komórkach, w których brak jest ekspresji
egrailed
i
hedgehog
.

Ekspresja genów wspomnianych kategorii
tworzy coraz to dokładniejszą mozaikę
sygnałów pozycyjnych,
które dostarczają każdej z komórek
informacji
precyzujących jej dokładną lokalizację w
zarodku.
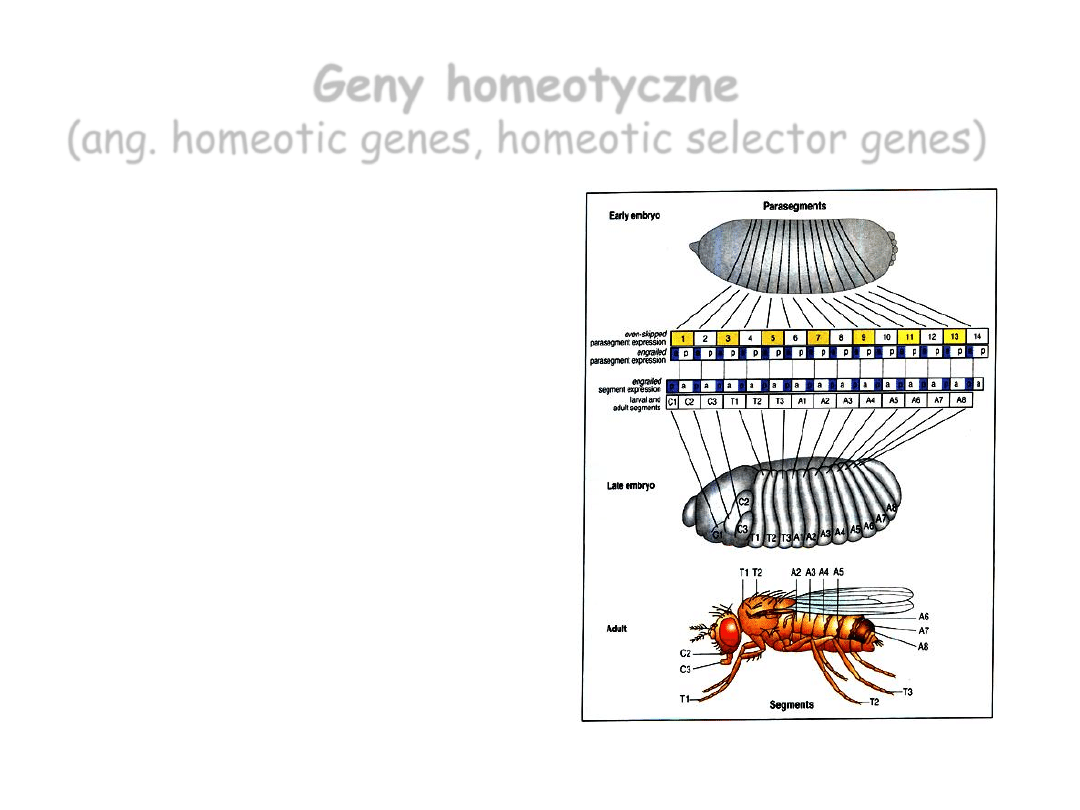
• Różnicują segmenty:
każdy segment
zdobywa własną
tożsamość, staje się
morfologicznie inny
od pozostałych.
Geny homeotyczne
(ang. homeotic genes, homeotic selector genes)

Geny
homeotyczne
zostały odkryte
przez
Edwarda Lewisa
(1978)
The Nobel Prize in Physiology or Medicine 1995

Lewis znalazł mutanta muszki owocowej
(
Bithorax
), który posiadał dwie pary
skrzydeł zamiast jednej pary.
Dodatkowa para wytworzyła się na
trzecim segmencie tułowiowym, ponieważ
mutacja (w genie
ultrabitorax
)
przekształciła trzeci segment tułowiowy
w segment drugi.
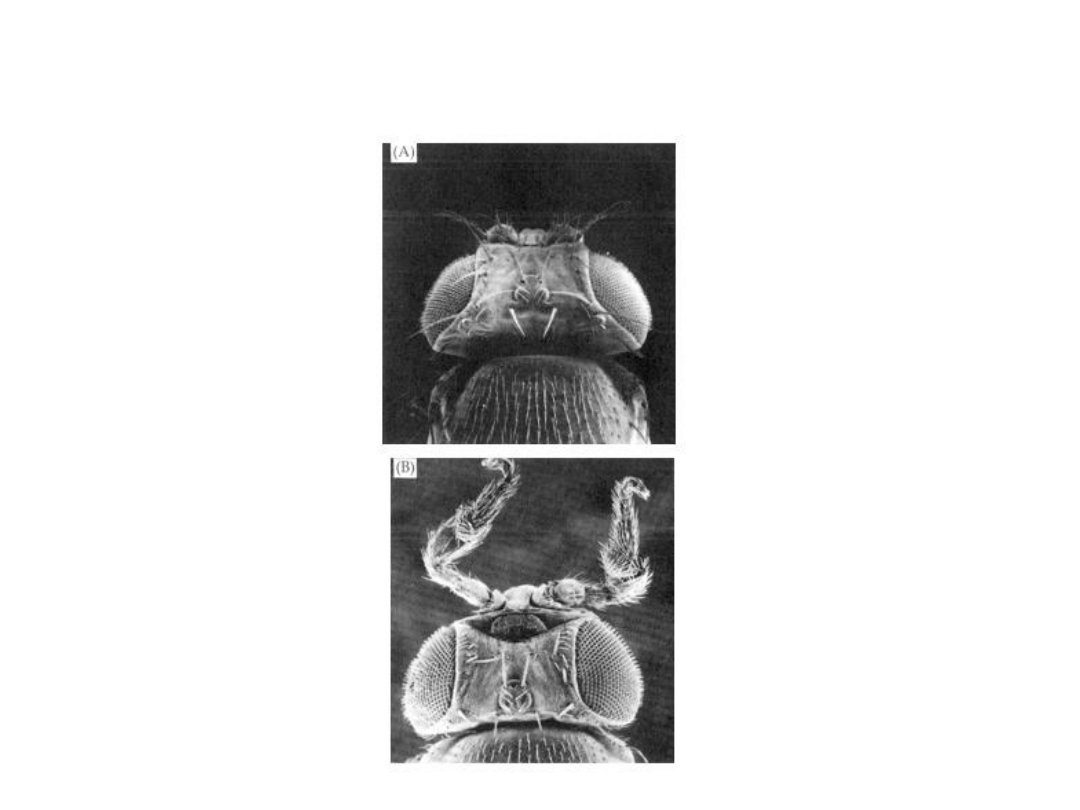
Mutacja
Antennapedia

Główne geny homeotyczne występują w
prawym ramieniu 3 chromosomu,
w postaci dwóch kompleksów genów
sprzężonych:
- Kompleks
Antennapedia
(
ANT-C
), wyznacza
różnice między segmentem głowowym a
trzema segmentami tułowia
- Kompleks
bithorax
(
BX-C
), wyznacza różnice
między segmentami tułowia a odwłoka.
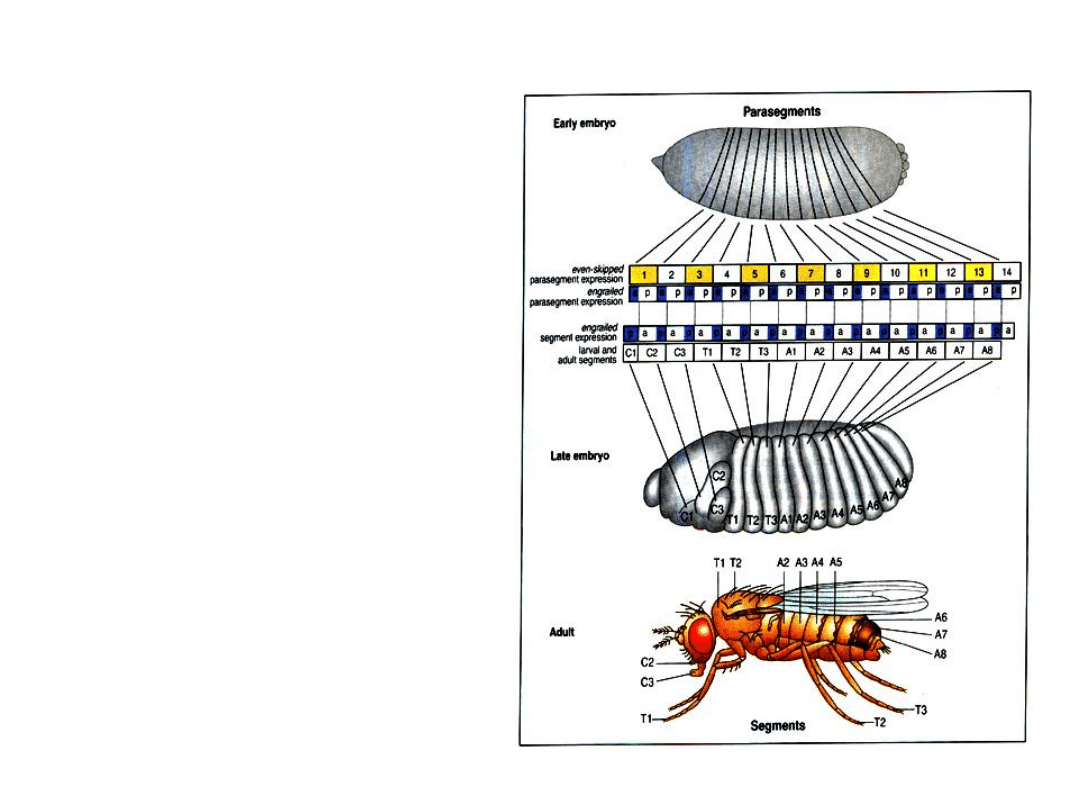
Geny obu kompleksów
ułożone są w
chromosomie w takiej
samej kolejności jak
parasegmenty, których
specjalizację
wyznaczają.

Geny homeotyczne cd.
• Ich ekspresją kierują białka (zestawy białek)
kodowane zarówno przez geny polarności jaja,
jak i przez geny segmentacji.
• Ekspresja g.h.rozpoczyna się bezpośrednio
przed celularyzacją blastodermy i początkowo
jest dość rozległa.
• W miarę jak aktywowane są kolejne grupy
genów segmentacji i przybywa kodowanych
przez te geny białek regulatorowych
ekspresja genów homeotycznych staje się
coraz bardziej precyzyjna.

• Klonowanie genów homeotycznych
ANT-C
i
BX-C
wykazało, że wszystkie zawierają
sekwencję o bardzo wysokim stopniu homologii
(o długości 180 nukleotydów)
.
• Sekwencja ta, odkryta w roku 1984 i nazwana
homeoboksem
(ang. homeobox), została potem
znaleziona również w genach polarności jaja, w
wielu genach segmentacji oraz w genach
kierujących rozwojem u różnych grup
zwierząt, także u ssaków.
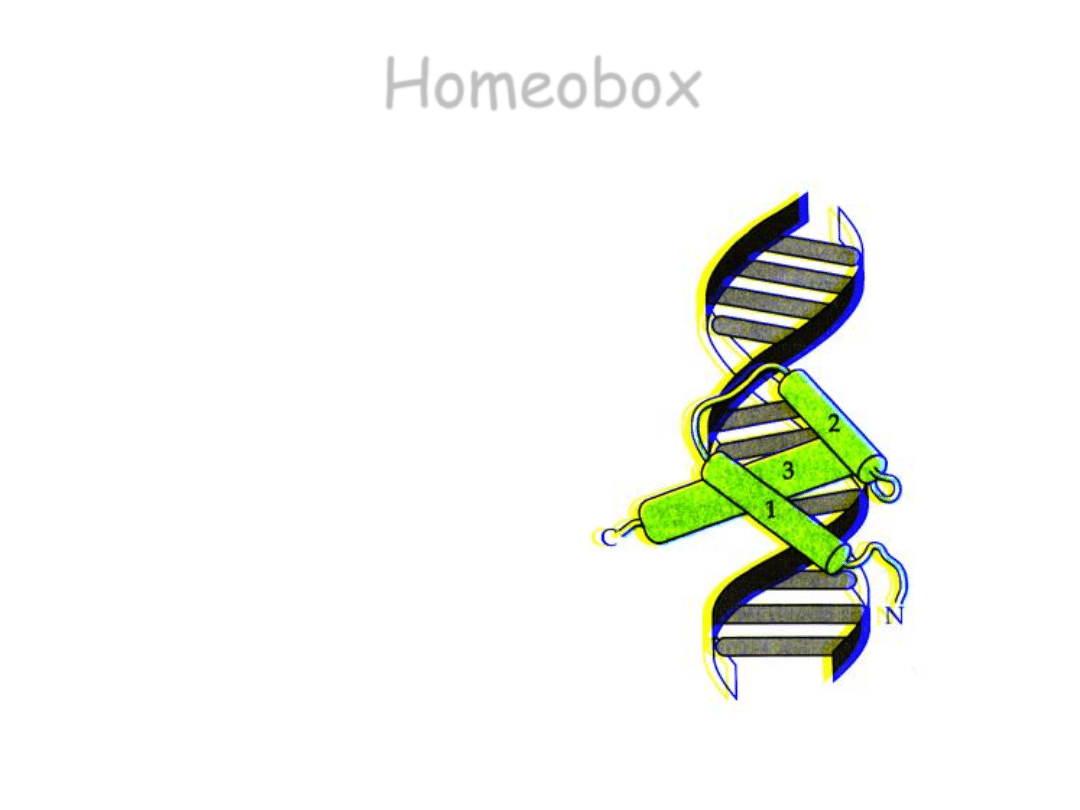
Homeobox
• Homeobox koduje domenę
białkową zwaną
homeodomeną
,
o dł. 60
aminokwasów, złożoną z
trzech fragmentów alfa-
helikalnych, tworzących
strukturę typu „heliks-skręt-
heliks”, typową dla czynników
transkrypcyjnych.
• Białka, które zawierają
homeodomenę biorą udział w
regulacji ekspresji zespołów
genów, które są ze sobą
funkcjonalnie powiązane.

Sekwencja homeoboksu występuje w
genach kierujących rozwojem u zwierząt
bezkręgowych i kręgowych.
• Posługując się sekwencją homeoboksu jako sondą do hybrydyzacji
DNA banków genomowych różnych zwierząt, stwierdzono
obecność homeoboksu w genach wielu bardzo odległych od siebie
ewolucyjnie grup:
• u nicienia
C.elegans
• u pierścienic
• u jeżowców
• u prymitywnych strunowców
• u kręgowców ( żaby, kury, myszy i człowieka)
• Geny zawierające homeoboks zostały dość dobrze poznane u
człowieka i u myszy, u których nazwano je genami
Hox
.
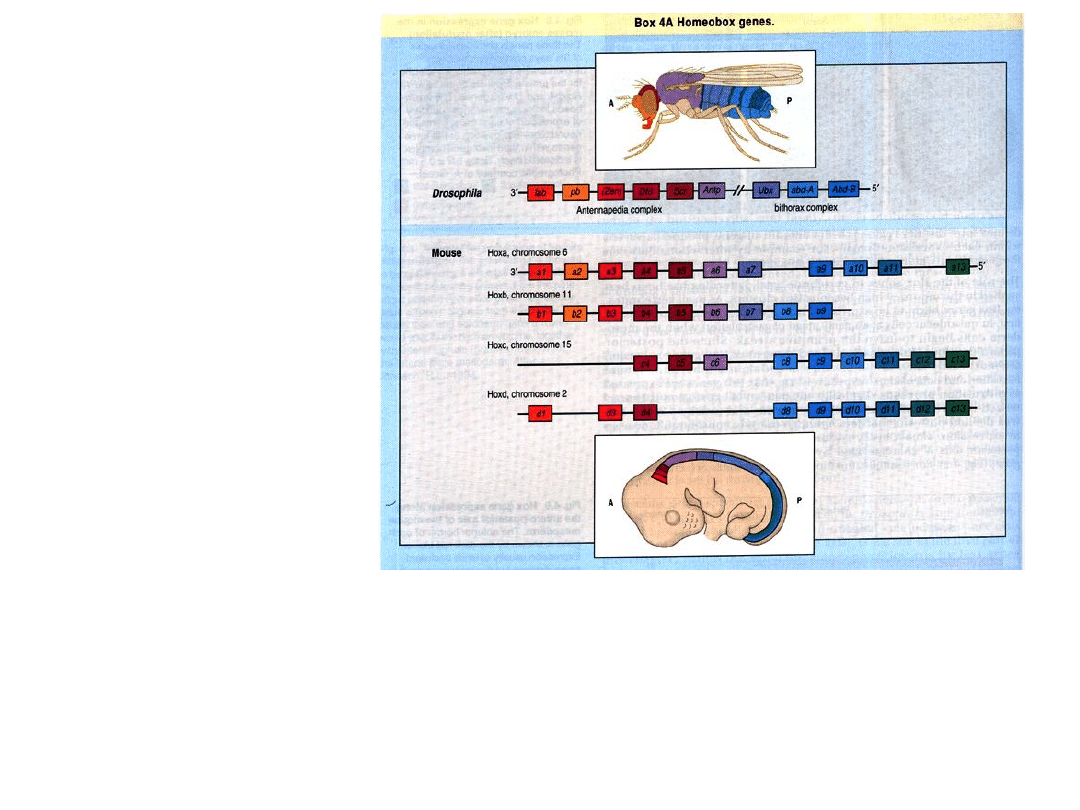
• Geny
Hox
występują w 4 grupach sprzężeń jako kompleksy
HOX
.
• Ułożenie genów w każdym z tych kompleksów wykazuje bardzo
duże podobieństwo do połączonych kompleksów
ANT-C
+
BX-C
u
Drosophila
: Najbardziej homologiczne w stosunku do siebie geny,
tworzące tzw. podrodziny, występują w tej samej kolejności.
Podobnie jak u
Drosophila
istnieje
ścisły związek między
kolejnością ułożenia
genów w chromosomie
a miejscem ich
aktywacji w osobniku.

• przednia granica ekspresji genu
(na przednio-tylnej osi ciała) zależy od pozycji genu w
kompleksie
• geny wykazujące ekspresję w tym samym regionie
pochodzą z tej samej podrodziny

Układ nerwowy
• Granica ekspresji poszczególnych genów
Hox
jest bardzo wyraźna
w układzie nerwowym zarodka myszy i odpowiada kolejności tych
genów w chromosomie.
• Geny
Hox
, których ekspresja pojawia się najpierw w narządach
osiowych (system nerwowy, zawiązki kręgów), wykazują ekspresję
również w narządach obwodowych, np. kończynach. Układowi
przednio-tylnemu w narządach osiowych odpowiada układ
proksymalno-dystalny w kończynach.
=> Ten sam system regulacyjny jest stosowany wielokrotnie, w
podobny sposób na różnych etapach rozwoju
Geny
Hox
zachowały swoje pierwotne funkcje:
• Po wprowadzeniu do
organizmu
Drosophila
genu
myszy
odpowiadającego
genowi
Antennapedia
i
spowodowaniu jego
nadmiernej ekspresji,
uzyskano muchy z odnóżami
zamiast czułków => podobnie
jak przy nadmiernej
ekspresji własnego genu.
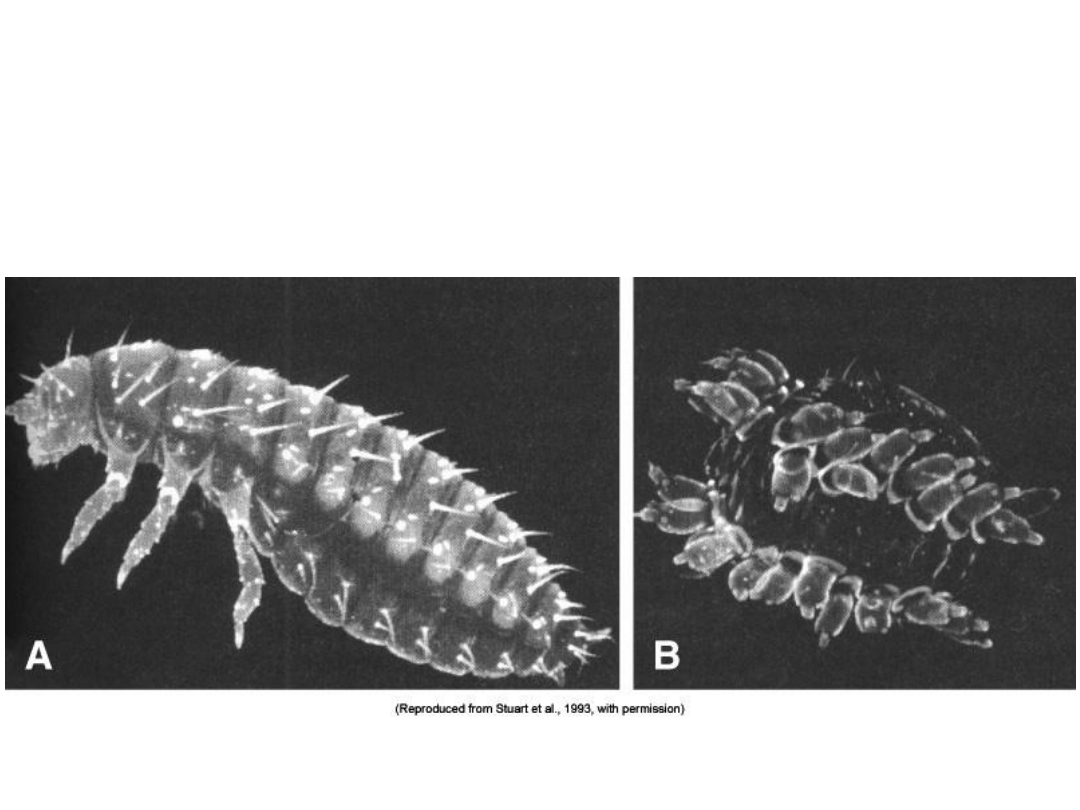
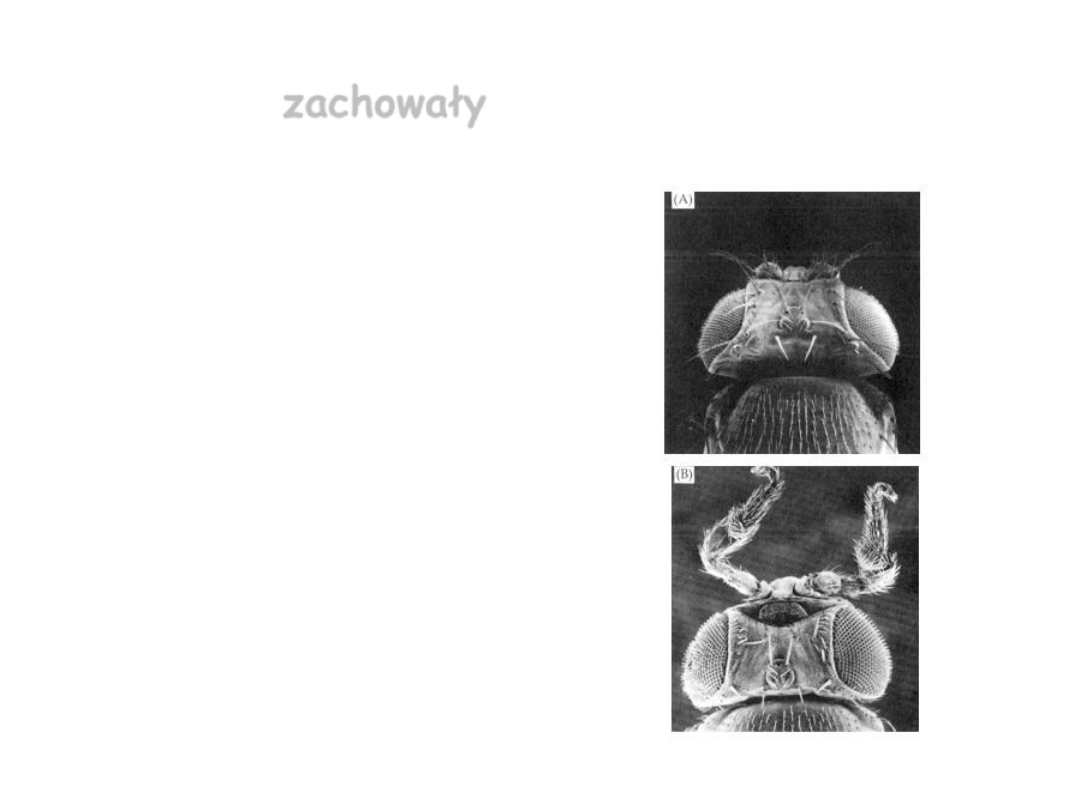
Eliminacja wszystkich genów
hox
u
Tribolium
–> wszystkie segmenty wyglądają tak samo
(Stuart i inni, 1993)
• A: normalny wygląd B: osobnik pozbawiony grupy genów
Hox
Wyszukiwarka
Podobne podstrony:
rozwoj ukladu nerwowego
organizacja i rozwój układu nerwowego
ROZWÓJ UKŁADU NERWOWEGO, Pedagogika
06 Rozwój układu nerwowego
ROZWÓJ UKŁADU NERWOWEGO, Anatomia Prawidłowa człowieka
rozwoj ukladu nerwowego
WADY ROZWOJOWE UKLADU NERWOWEGO
Wady rozwojowe układu nerwowego
Rozwój układu nerwowego
Rozwój układu nerwowego (2)
1 Rozwój ukladu nerwowego 2010 11id 8683 pptx
SPECYFIKA CHORÓB UKŁADU NERWOWEGO OKRESU ROZWOJOWEGO
Elementy patologii układu nerwowego wykiład 1, Pedagogika, Wczesne wspomaganie rozwoju, wczesne wspo
Karta przedmiotu A M-B-1, Pedagogika, Wczesne wspomaganie rozwoju, wczesne wspomaganie rozwoju, seme
SPECYFIKA CHORÓB UKŁADU NERWOWEGO OKRESU ROZWOJOWEGO
Znaczenie układu nerwowego dla rozwoju dziecka oraz procesu uczenia się Specyficzne trudności w ucze
Choroby układu nerwowego ppt
więcej podobnych podstron