
1
GENETYKA
Cykl wykładów 1
dr hab. Maria Wedzony 2007
1. Historia genetyki
2. Budowa i replikacja DNA
3. Niektóre aspekty podziałów komórkowych
2
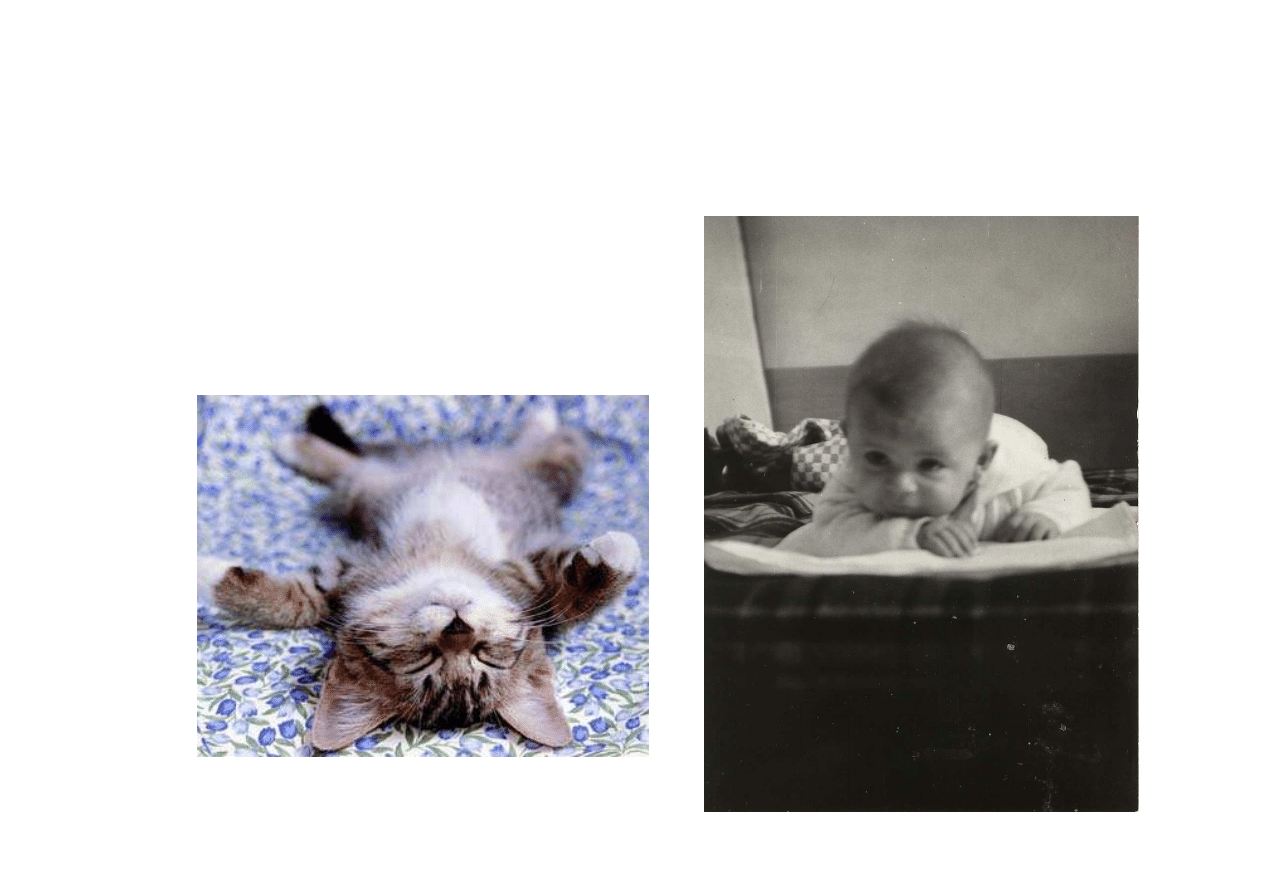
Od niepamiętnych czasów obserwowano
podobieństwo potomstwa do rodziców.
Człowiek rodzi człowieka…
Kot rodzi kota…

3
Ale dlaczego dzieci nie są identyczne z rodzicami?
Jak to się dzieje, że kobieta rodzi także chłopców?
Skąd u dzieci pojawiają się cechy nieobecne u żadnego z rodziców?

4
Jaka jest rola
osobnika
męskiego w
rozmnażaniu, w
przekazywaniu
cech na
potomstwo?

5
Jeszcze więcej problemów
nastręczały organizmy niższe:
Aż do XIX wieku powszechne
było przekonanie, że muchy
rodzą się z brudu, a żaby z
błota. Dopiero wykrycie
mikroskopu podważyło te
teorie.

6
Zanim zrozumiano na czym
polega dziedziczenie cech
już wykorzystywano reguły
dziedziczenia w praktyce:
w hodowli roślin i zwierząt.
Krzyżując zwierzęta o
określonych cechach i
blisko spokrewnione
otrzymano rasy hodowlane
zwierząt domowych i
odmiany roślin uprawnych.
Hodowcy posługiwali się
intuicją i obserwacją.

7
Gregor Johann Mendel: ur. 20 lipca 1822 w Hynczycach koło
Nowego Jiczyna, wówczas Cesarstwo Austro-W
ę
gierskie – zm. 6
stycznia w Brnie Morawskim, Cesarstwo A-W)
Aagustianin, prekursor genetyki.
Klasztor w Brnie na Morawach
Grzegorz Mendel

8
Grzegorz Mendel
W 1866 roku opublikował pracę o
dziedziczeniu niektórych cech grochu
zawierającą do dziś obowiązujące
fundamentalne prawa genetyki. Nie on
pierwszy badał dziedziczenie. Jak mu się
udało?
•Znał dobrze matematykę, a w tym – statystykę
=> pracował z dużymi grupami osobników (po
kilka tysięcy w jednej próbie)
•Krzyżowania rozpoczynał dopiero wtedy, gdy
był pewien, że materiał wykazuje te same cechy
przez kilka pokoleń.
•Biologia kwitnienia grochu sprzyjała
doświadczeniom.
•Potomstwo krzyżowań badał przez kilka
pokoleń prowadząc bardzo staranne obliczenia.
•Kiedy badał dziedziczenie wybranych cech nie
zwracał uwagi na pozostałe cechy organizmu.

9
Element szczęścia:
•
Wybrał łatwe do zaobserwowania cechy, determinowane jednogenowo i każda o
dwóch allelach o prostych stosunkach dominacji.
•
Wybrał cechy determinowane genami leżącymi na różnych chromosomach.
Mikroskop z czasów Mendla i projekt oran
ż
erii na dziedzi
ń
cu klasztoru w Brnie,
autorstwa Mendla, cz
ęś
ciowo zrealizowany kiedy był przeorem.

10
Z tego względu pod
koniec życia Mendel
zaczął badać wiesiołek
Oenothera biennis
.
Badania te
doprowadziły go do
głębokiej depresji, gdyż
żadne z wcześniej
odkrytych praw się nie
sprawdzało!
Współcześni nie zrozumieli teorii Mendla i nie uwierzyli w jego
obliczenia – biolodzy w tamtych czasach przeważnie nie znali
matematyki na wystarczającym poziomie by przemówiły do nich
matematyczne argumenty. Zarzucano mu ponadto, że wszystkie
badania przeprowadził na jednej roślinie, więc jego prawa nie mogą
mieć charakteru uniwersalnego.
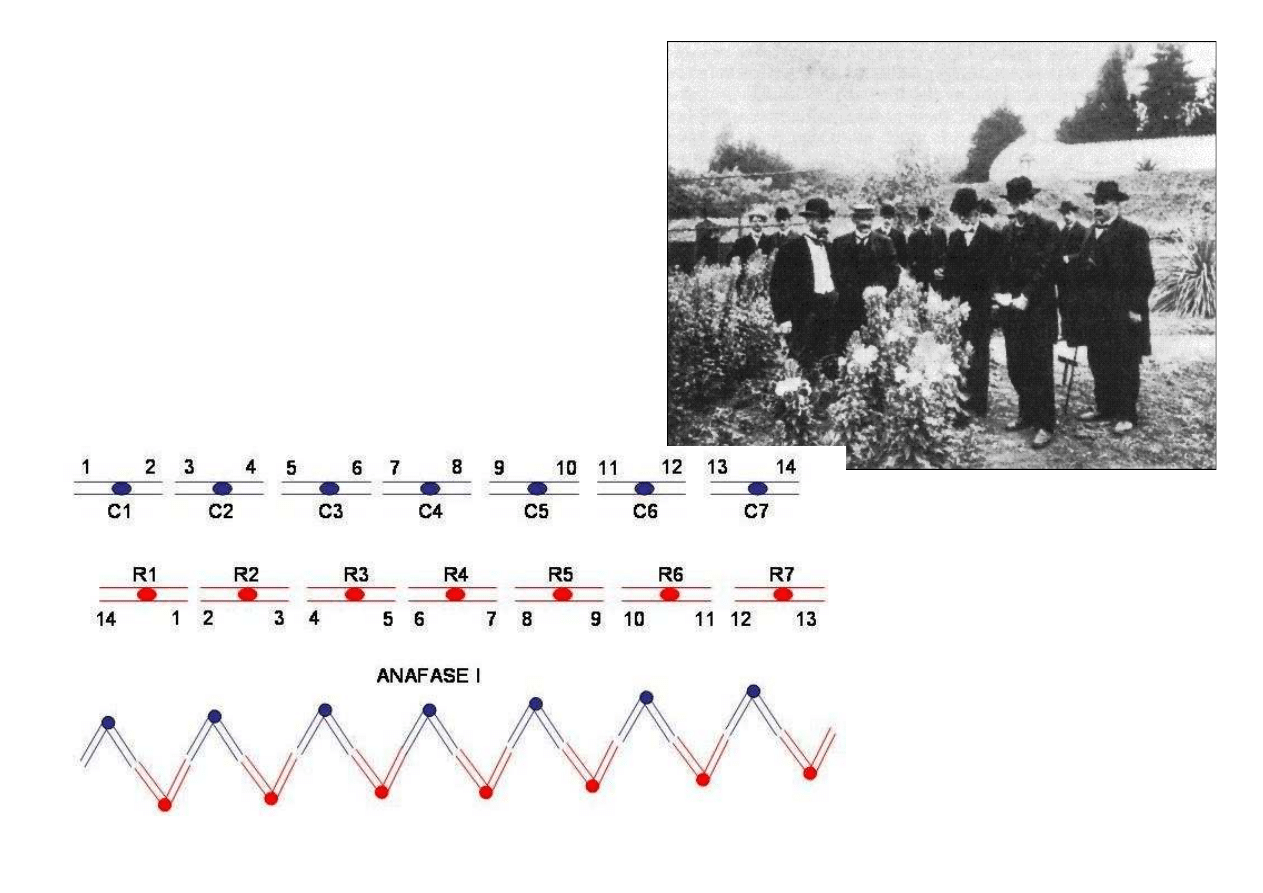
11
Dziś wiemy że:
Wiesiołek ma szereg translokacji w
swoich chromosomach i koniugują
one wszystkie w kształcie pierścienia
– nie ma niezależnego dziedziczenia
cech.
Obok: Hugo de Vries wraz z kolegami
oglądają rośliny wiesiołka w ogrodzie
botanicznym: początek XX wieku –
już wiedzą, dlaczego Mendlowi nie
udało się potwierdzić teorii na tej
roślinie.
Z lewej: Koniugacja u
wiesiołka. Chromosomy
rodzicielskie w mejozie tworz
ą
poliwalent w kształcie
pier
ś
cienia. Chromosomy
ka
ż
dego rodzica zawsze
rozchodz
ą
si
ę
do tego samego
bieguna: brak niezale
ż
nej
segregacji chromosomów, a
wi
ę
c i genów na nich
zlokalizowanych.
12
Kalendarium odkry
ć
dotycz
ą
cych dziedziczenia
Morgan i jego
grupa (Nobel w
1933 r.)
Geny
le
żą
linearnie na chromosomach
Mutacje
s
ą
zmianami fizycznymi w genach
Rekombinacje
genów zachodz
ą
w mejozie
1913
1927
1931
De Vries
Tschermack
Correns
Boveri
Sutton
Johansen
Oficjalne przyj
ę
cie praw Mendla
Teoria mutacji (de Vries)
Chromosomy s
ą
dziedziczone
Johansen – termin „gen”
Geny le
żą
na chromosomach
1900
1901
1903
1909
1910
Mendel
Jednostki dziedziczenia maj
ą
podło
ż
e materialne
1865
13
Avery
McLeod
MacCarty
Ostateczne potwierdzenie,
ż
e DNA jest materiałem
genetycznym
1944
Beadle
Tatum
Mutacje pokarmowe u Neurospora crassa:
teoria 1 gen = 1 enzym
1941
Casperson
Chromosomy zawieraj
ą
DNA
1936
Griffith
Hipoteza,
ż
e cechy dziedziczne zwi
ą
zane s
ą
z DNA –
transformacja bakterii Diplococcus
1928
Miescher
Wyodr
ę
bnienie kwasów nukleinowych ze spermy
łososia
1869
Kalendarium odkry
ć
dotycz
ą
cych dziedziczenia c.d.
14
Watson i
Crick
Model przestrzenny budowy DNA (Nobel 1962 r.)
1953
Chargraff
Adenina i tymina, oraz Cytozyna i Guanina
wyst
ę
puj
ą
w DNA zawsze w tych samych ilo
ś
ciach,
za to proporcje A-T i C-G s
ą
charakterystyczne dla
gatunku
1953
Hershey i
Chase
Podczas infekcji komórki bakteryjnej przez
bakteriofaga do wn
ę
trza wnika wył
ą
cznie DNA
1952
Rosalinda
Franklin
Zdj
ę
cia rentgenowskie krystalicznego DNA:
prawdopodobnie spirala?
1944
Kalendarium odkry
ć
dotycz
ą
cych dziedziczenia c.d.
15
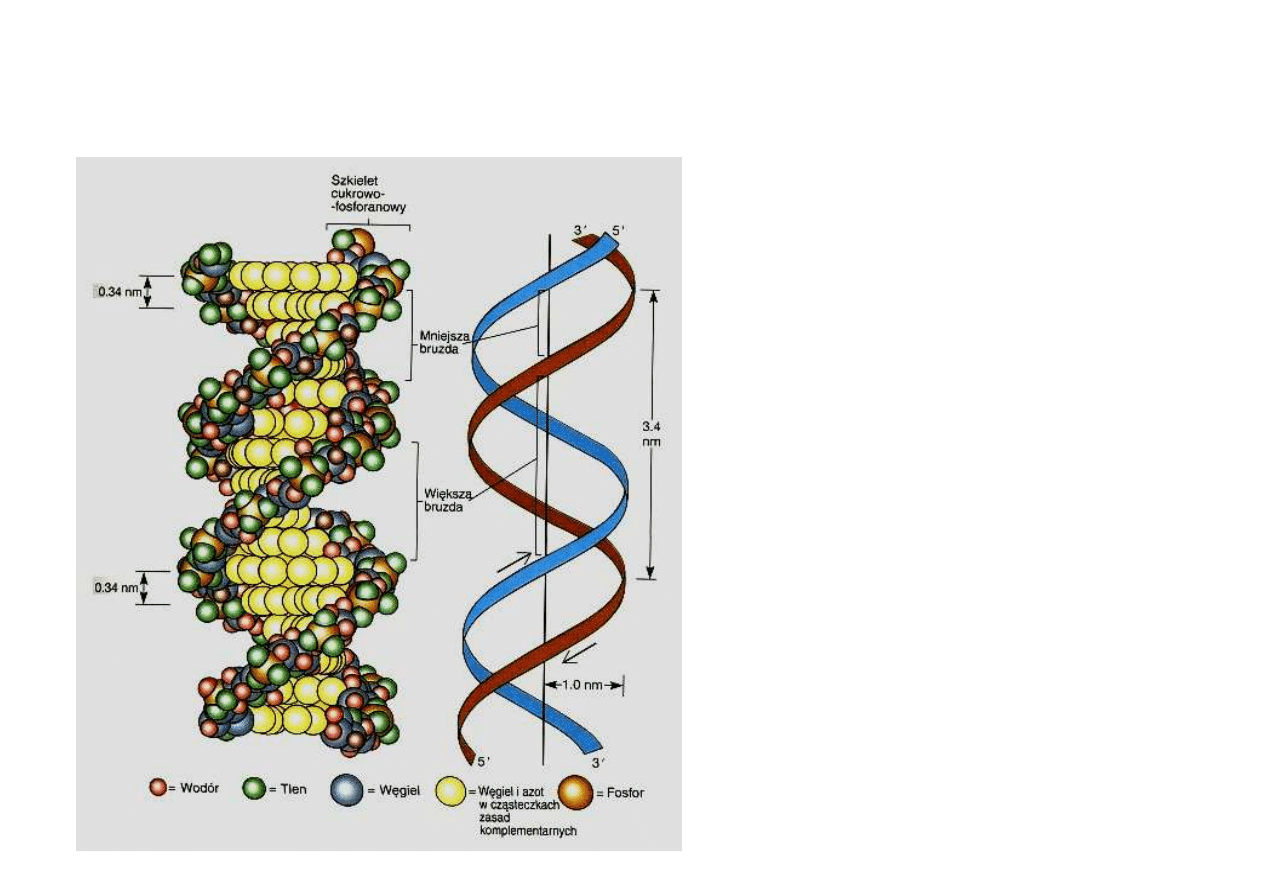
Co dzisiaj wiemy o DNA
DNA jest polimerem, który
swoje wła
ś
ciwo
ś
ci
zawdzi
ę
cza
specyficznej budowie
zasad, z których jest
zbudowany.
Specyficzna lokalna
konfiguracja
nukleotydów sprawia,
ż
e podwójna helisa nie
jest tak regularna w
budowie jak wyobra
ż
ali
sobie Watson i Crick.
16
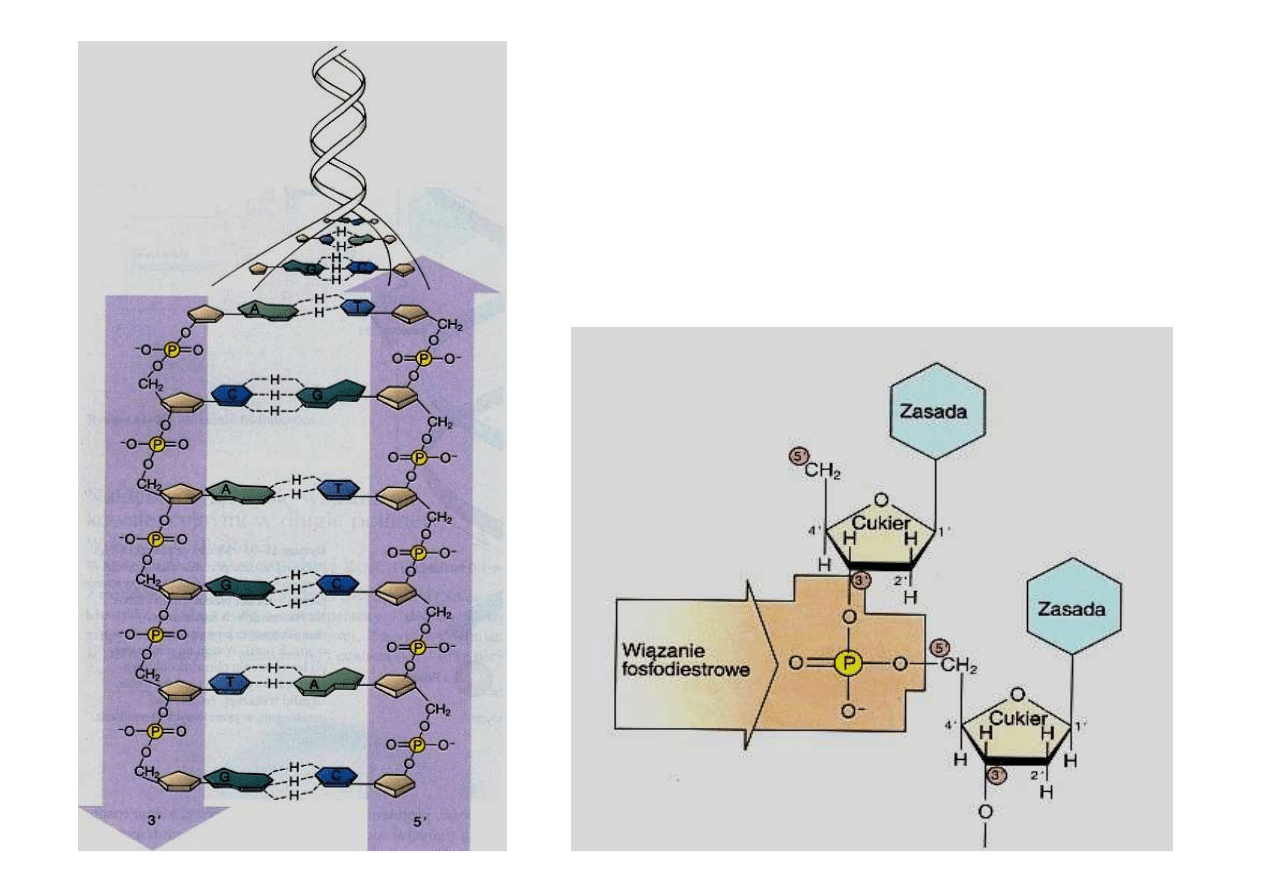
W dwuniciowym DNA
pomi
ę
dzy adenin
ą
i tymina
oraz pomi
ę
dzy guanin
ą
i
cytozyn
ą
tworz
ą
si
ę
wi
ą
zania wodorowe
.
17
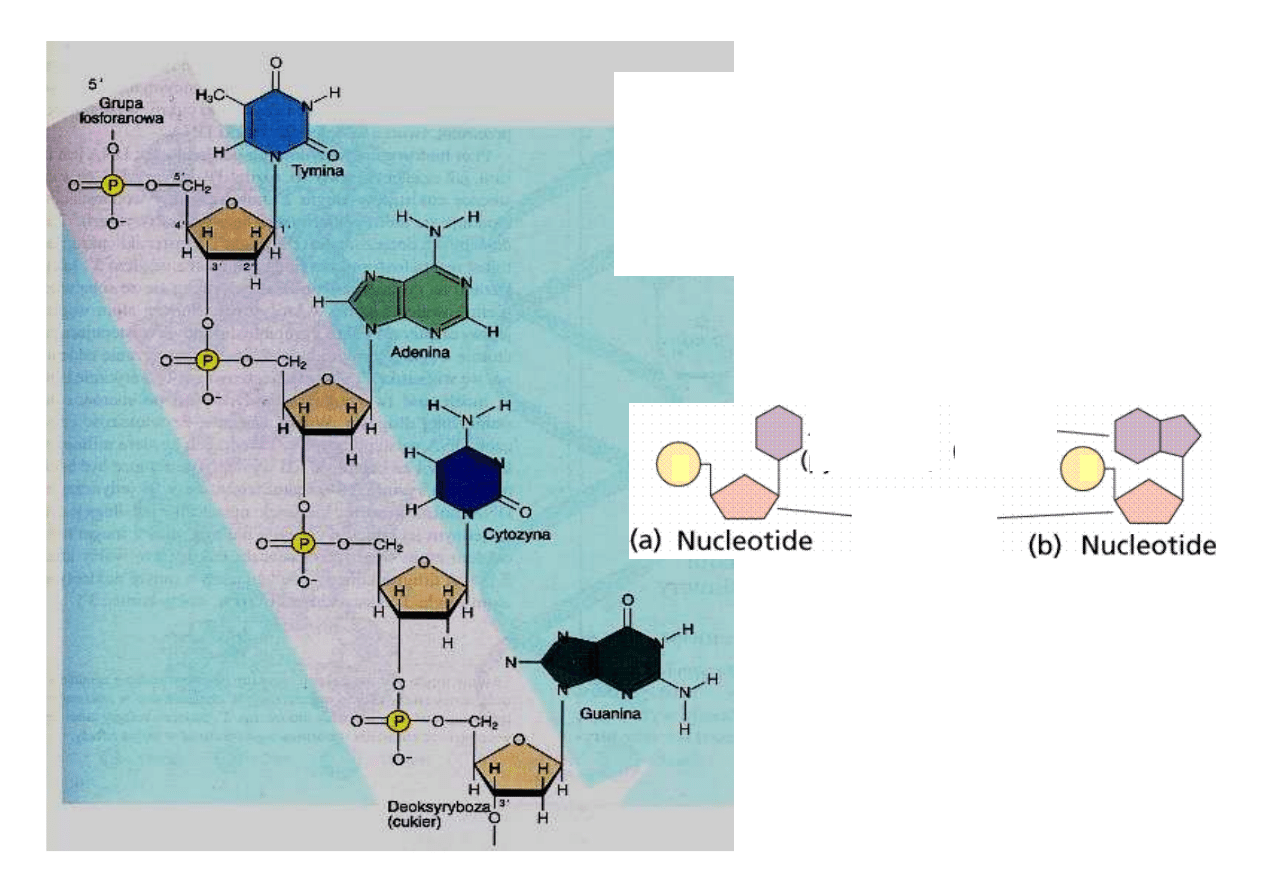
Z jednej strony ła
ń
cucha wolnym
miejscem do wi
ą
zania chemicznego
jest w
ę
giel 5’ pier
ś
cienia rybozy, a na
drugim ko
ń
cu ła
ń
cucha jest to w
ę
giel
3’. Odpowiednio ko
ń
ce pojedynczego
ła
ń
cucha DNA nazywamy 5’ i 3’
P
P
Zasada
pirymidyno
wa
Zasada
purynowa
Fosforan
rybozy
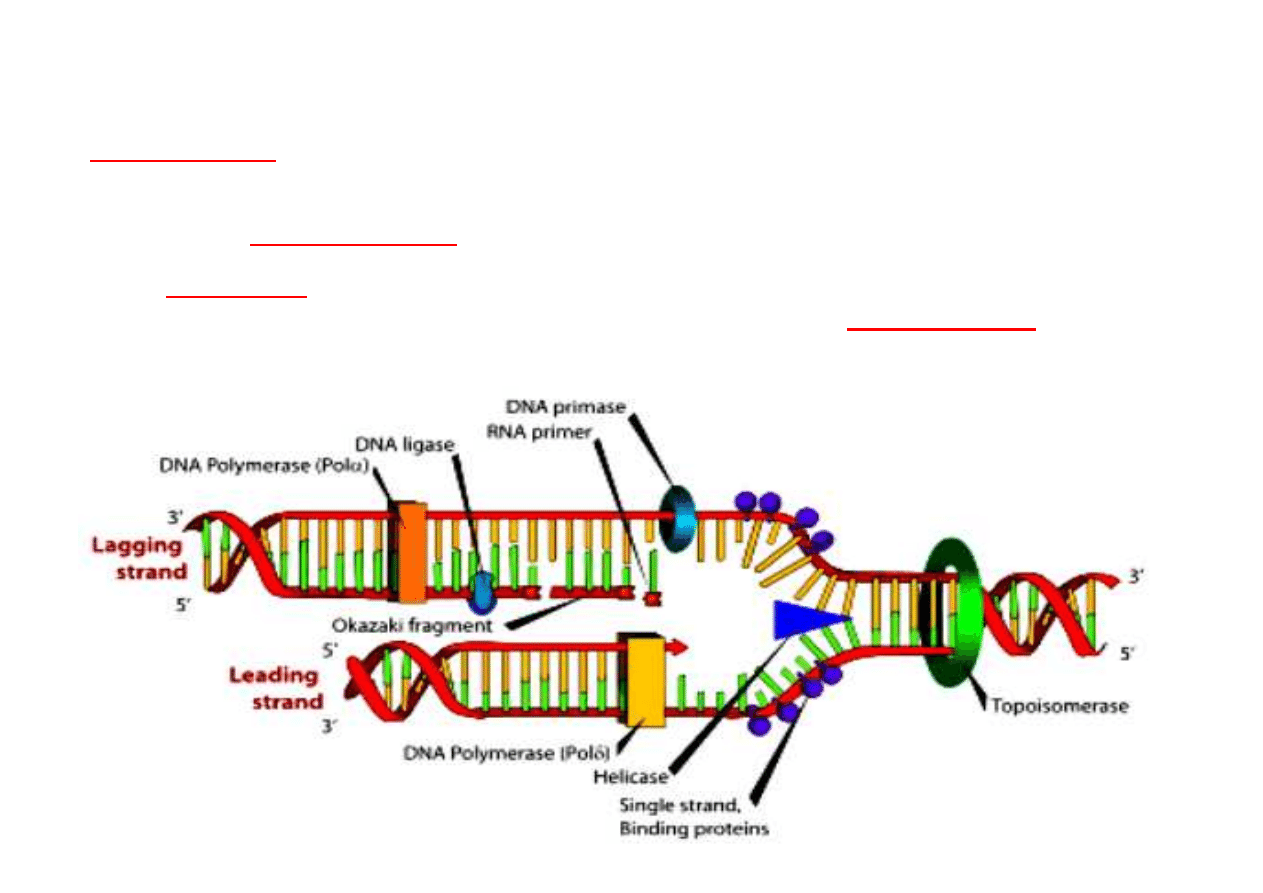
18
REPLIKACJA DNA
Aby mogła zaj
ść
replikacja podwójna helisa musi by
ć
rozpleciona za pomoc
ą
helikazy DNA.
Do rozł
ą
czaj
ą
cego si
ę
miejsca przył
ą
czaj
ą
si
ę
białka destabilizuj
ą
ce
heliks (single strand binding proteins)
i umo
ż
liwiaj
ą
rozpocz
ę
cie replikacji.
Tylko jedna ni
ć
(
ni
ć
prowadz
ą
ca – leading strand
) syntetyzowana jest w sposób
ci
ą
gły przez
polimeraz
ę
DNA
. Druga (
ni
ć
opó
ź
niona – lagging strand
) syntetyzowana
jest we fragmentach 100-1000 nukleotydów (
fragmenty Okazaki
), które sklejane s
ą
przez
ligaz
ę
DNA
.
Napi
ę
cia powstaj
ą
ce w nici na skutek rozplatania likwiduj
ą
topoizomerazy
rozcinaj
ą
c co jaki
ś
czas obie nici helisy umo
ż
liwiaj
ą
c im rozkr
ę
cenie si
ę
, a
nast
ę
pnie ł
ą
cz
ą
c precyzyjnie rozci
ę
te ko
ń
ce
.
19
DNA syntetyzowany jest zawsze w kierunku od 5' do 3‘. Do rozpocz
ę
cia
syntezy DNA potrzebny jest krótki fragment RNA zwany starterem
(primerem) syntetyzowanym przez kompleks białkowy nazywany
primeosomem (replisomem).
Polimeraza DNA dobudowuje nowe nukleotydy do ko
ń
ca 3’
starterowego RNA. Startery przył
ą
czaj
ą
si
ę
do macierzystej nici DNA w
ś
ci
ś
le okre
ś
lonych miejscach nazywanych „miejscami rozpocz
ę
cia
replikacji” z angielskiego ORI. Po spełnieniu swojej roli starterowy RNA
zostaje wyci
ę
ty i rozło
ż
ony.
Replisom czyli primosom
/ prymosom (aparat replikacyjny) – grupa białek
rozpoczynaj
ą
cych i kontynuuj
ą
cych replikacj
ę
DNA.
Jest to replikacyjny kompleks białkowy obejmuj
ą
cy wiele białek m.in. s
ą
to: polimeraza DNA III, primaza, białko DnaA (inicjatorowe), białko DnaB
(helikaza), białko DnaC. Rozwija on widełki replikacyjne podczas
replikacji.
Szybko
ść
replikacji replisomu wynosi kilkaset bp/s. Efektywna szybko
ść
replikacji DNA wynosi od 0 do 1000 bp/s w optymalnych warunkach. In
vivo u Eucaryota proces przebiega
ś
rednio znacznie wolniej i zale
ż
y od
fazy rozwojowej komórki.
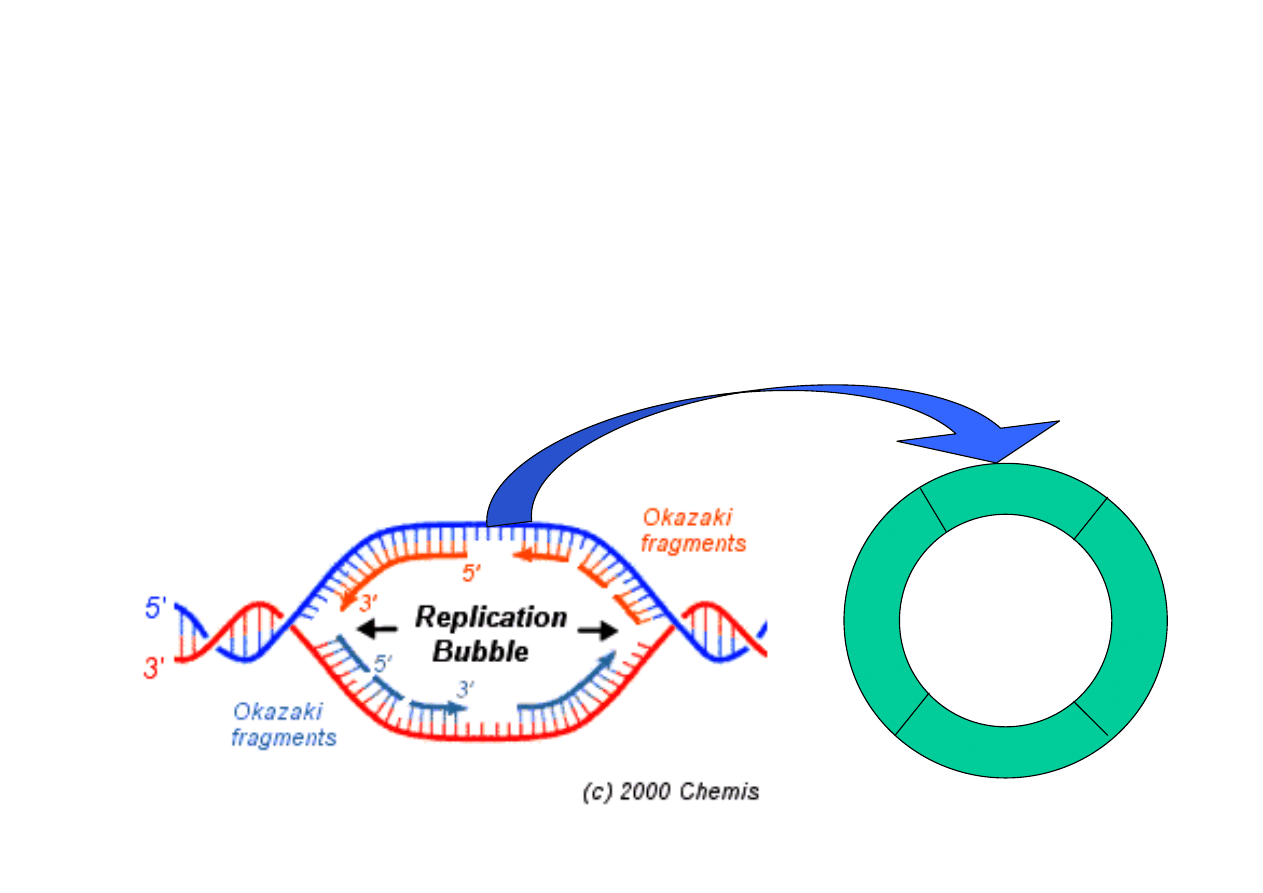
20
Miejsce rozł
ą
czenia si
ę
nici i post
ę
pu replikacji nazywamy
widełkami
replikacyjnymi
. Przeciwstawne widełki tworz
ą
b
ą
bel replikacyjny
(replication bubble). W fazie syntezy cyklu podziałowego tworz
ą
si
ę
setki/tysi
ą
ce b
ą
bli replikacyjnych. Replikacja trwa do momentu, kiedy
spotkaj
ą
si
ę
ze sob
ą
widełki replikacyjne zd
ąż
aj
ą
ce w przeciwnych
kierunkach, a ligaza DNA poł
ą
czy oba ko
ń
ce nowo utworzonych nici.
W rzeczywisto
ś
ci proces jest znacznie bardziej skomplikowany, gdy
ż
ł
ą
czy
si
ę
z rozpadem i odbudow
ą
nukleosomów na obu niciach, tworzeniem
struktur DNA wy
ż
szego rz
ę
du. W procesie bior
ą
równie
ż
udział enzymy
naprawcze, sprawdzaj
ą
ce poprawno
ść
replikacji i naprawiaj
ą
ce drobne
bł
ę
dy kopiowania.
Cykl
podziałowy
mitoza
synteza
G1
G2
Semi-konserwatywna
replikacja DNA
Wyszukiwarka
Podobne podstrony:
wykłady z genetyki i parazytologii 2007-2008 część 2 - paraz, farmacja, parazytologia
wykłady z genetyki i parazytologii 2007-2008 część 1 - syste, farmacja, parazytologia
Gleby RW w1 2007
Genetyka – egzamin 2007
wykłady z genetyki i parazytologii 2007-2008 część 3 - genet, farmacja, parazytologia
wykłady z genetyki i parazytologii 2007-2008 część 2 - paraz, farmacja, parazytologia
2007, 5 (oniszczenko), Elementy genetyki zachowania Ćw
Ćwiczenia studenci - 02.10.2007, Ogrodnictwo UP Wrocław, Semestr IV, Genetyka
28.10.2007, biologia z genetyką
W1 1 Wprowadzenie 2007
AM1 2007 W1
Test podstawy genetyki 2007, Biologia - testy liceum
Genetyka 2007.2008 zaliczenie I (termin II), genetyka, testy, testy genetyka kolokwium nr 1
pytania genetyka sesja wiosna 2007, Genetyka, testy, lekarskie, ustny
genetyka klasyczna, W1-2R05, 1
2007 Godlewska Genetyka i psychiatria
więcej podobnych podstron