1) Przedstaw graficznie i omów potencjały czynnościowe tkanki roboczej i bodźcotwórczej serca oraz mechanizmy jonowe leżące u ich podstawy.
Potencjały czynnościowe w sercu
Bodziec działający na spolaryzowaną komórkę mięśnia sercowego (prawidłowo z węzła zatokowo-przedsionkowego) zmienia przepuszczalność błony dla jonów sodu, które dostając się do wnętrza komórki, zmniejszają ujemny potencjał do wartości ok. -65 mV (potencjał progowy).
Przekroczenie potencjału progowego jest czynnikiem wyzwalającym otwarcie kanałów sodowych.
Dochodzi wówczas do gwałtownego napływu jonów sodu do wnętrza komórki, w wyniku czego następuje szybka i całkowita depolaryzacja.
Przy wartości -40 mV otwierają się z lekkim opóźnieniem kanały wapniowe.
W powstającym potencjale czynnościowym wyróżniamy pięć faz:
faza 0 (szybka depolaryzacja) - zależy od szybkiego dośrodkowego prądu sodowego
faza 1 (wstępna szybka repolaryzacja) - dośrodkowy prąd chlorkowy i odśrodkowy prąd potasowy
faza 2 (powolna repolaryzacja) - tzw. faza plateau (stabilizacja potencjału równowagą pomiędzy dośrodkowym prądem wapniowo-sodowym a odśrodkowym prądem potasowym)
faza 3 (szybka repolaryzacja) - przewaga odśrodkowego prądu potasowego nad wygasającym dośrodkowym prądem wapniowo-sodowym
faza 4 (polaryzacja) - faza spoczynku, polaryzacji
Potencjał czynnościowy w mięśniu sercowym:
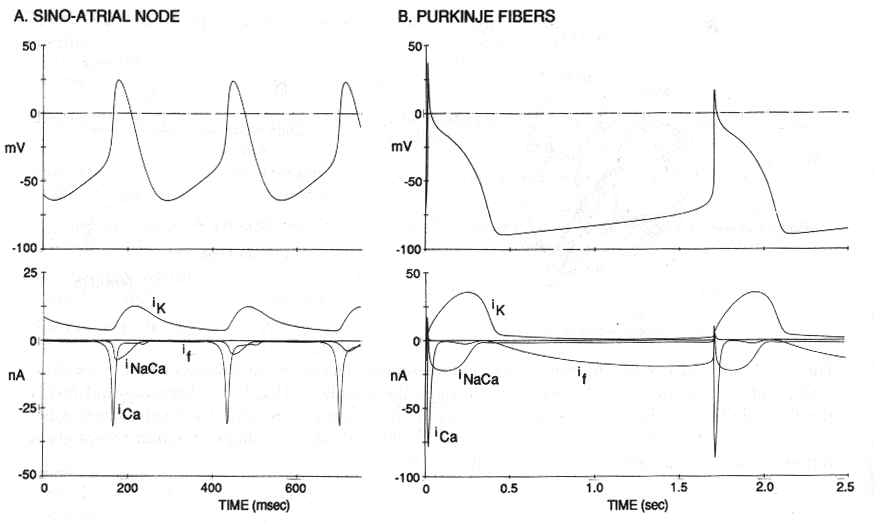
Mechanizm powstawania potencjału czynnościowego w mięśniu sercowym:
Powolna depolaryzacja - potencjał rozrusznikowy (spadek IK, wolny wzrost ICa)
Szybki wzrost - nagły wzrost ICa
Powolny spadek - spadek INaCa i wzrost IK
Wyjście z hiperpolaryzacji - wzrost If (prąd Na i K aktywowany hiperpolaryzacją)
Komórki rozrusznikowe serca mają zdolność do tzw. spontanicznej powolnej depolaryzacji.
W jaki sposób kończy się potencjał czynnościowy komórki mięśnia sercowego?
- Przepuszczalność dla jonów wapniowych stopniowo zmniejsza się podczas utrzymującej się depolaryzacji.
- To zmniejszenie może być następstwem stopniowego zwiększenia wewnątrzkomórkowego stężenia wapnia w wyniku napływu jonów wapniowych do wnętrza komórki przez otwarte kanały wapniowe.
- Nagromadzone w komórce jony wapniowe zaczynają oddziaływać bezpośrednio i pośrednio na kanały wapniowe, powodując ich zamknięcie.
- Przepuszczalność błony komórkowej dla jonów potasu zwiększa się, co przesuwa potencjał błonowy w kierunku potencjału równowagi dla jonów potasu i repolaryzuje komórkę mięśnia sercowego.
- Wydaje się, że zwiększenie przepuszczalności dla jonów potasu jest częściowo związane z działaniem kanałów potasowych zależnych od napięcia, które otwierają się pod wpływem depolaryzacji w trakcie trwania potencjału czynnościowego (podobnie jak bramki n kanałów błonowych neuronu).
- Kanały potasowe aktywowane jonami wapnia mogą również uczestniczyć w opóźnionym zwiększeniu przepuszczalności dla jonów potasu.
Jedną z czynnościowych konsekwencji przedłużonego czasu trwania potencjału czynnościowego w mięśniu sercowym jest fakt, ze czas trwania potencjału czynnościowego wpływa na czas trwania skurczu mięśnia sercowego.
- W mięśniu sercowym tylko początek skurczu jest zależny od wapnia uwalnianego z siateczki sarkoplazmatycznej; skurcz podtrzymwany jest przez jony wapniowe napływające do komórki przez błonę komórkową w fazie plateau potencjału czynnościowego.
- Dlatego też można zmienić czas trwania skurczu komórki mięśnia sercowego, zmieniając czas trwania potencjału czynnościowego we włóknie mięśniowym.
- Zmiany potencjału czynnościowego stanowią istotny mechanizm modulujący pracę serca jako pompy.
Funkcje węzła SA i AV
Węzeł SA:
*W prawidłowo funkcjonującym sercu częstość pracy serca kontrolują komórki rozrusznikowe, zlokalizowane w węźle zatokowo-przedsionkowym (SA), zlokalizowanym w górnej części prawego przedsionka.
- Potencjał czynnościowy komórek tego węzła różni się od potencjału czynnościowego innych komórek serca.
- W węźle przedsionkowo-komorowym większą rolę odgrywają kanały wapniowe niż sodowe w powstawaniu potencjału czynnościowego i utrzymywaniu fazy depolaryzacji.
*W pracującym w spoczynku zdrowym sercu człowieka komórki węzła zatokowego generują potencjały czynnościowe z częstością ok. 70 na minutę.
*Te potencjały czynnościowe rozchodzą się na drodze elektrycznej w przedsionkach, powodując skurcz przedsionków.
Węzeł AV:
*Patrząc pod kątem przewodnictwa elektrycznego możemy powiedzieć, że serce zawiera dwie izolowane podjednostki.
- Dwa przedsionki stanowią jedną podjednostkę, a dwie komory drugą.
*Połączenia elektryczne pomiędzy tymi dwiema jednostkami dokonują się za pośrednictwem wyspecjalizowanych komórek mięśniowych, tworzących węzeł przedsionkowo--komorowy (AV).
- Pobudzenie z przedsionków przechodzi przez węzeł przedsionkowo-komorowy.
- Komórki węzła przedsionkowo-komorowego mają małą średnicę w porównaniu z komórkami mięśnia sercowego.
- Zwolnienie przewodzenia w węźle przedsionkowo-komorowym wpływa na opóźnienie skurczu komór w stosunku do skurczu przedsionków.
3) Scharakteryzuj poszczególne załamki, odcinki i odstępy w prawidłowym EKG
Charakterystyka EKG - elektrokardiogram:
Na wykresie EKG analizuje się:
linię izoelektryczną - linia pozioma zarejestrowana w czasie, gdy w sercu nie stwierdza się żadnych pobudzeń (aktywności). Najłatwiej wyznaczyć ją według odcinka PQ. Stanowi ona punkt odniesienia poniższych zmian
załamki - wychylenia od linii izoelektrycznej (dodatni, gdy wychylony w górę; ujemny, gdy wychylony w dół)
odcinki - czas trwania linii izoelektrycznej pomiędzy załamkami
odstępy - łączny czas trwania odcinków i sąsiadującego załamka
Załamki
załamek P - jest wyrazem depolaryzacji mięśnia przedsionków (dodatni we wszystkich 11 odprowadzeniach, poza aVR, tamże ujemny)
zespół QRS - odpowiada depolaryzacji mięśnia komór
załamek T - odpowiada repolaryzacji komór
czasem też załamek U
Odcinki
odcinek PQ - wyraża czas przewodzenia depolaryzacji przez węzeł przedsionkowo-komorowy (AV)
odcinek ST - okres depolaryzacji komór
Odstępy
odstęp PQ - wyraża czas przewodzenia depolaryzacji od węzła zatokowo-przedsionkowego do węzeł przedsionkowo-komorowy (SA -> AV)
odstęp ST - wyraża czas wolnej i szybkiej repolaryzacji mięśnia komór (2 i 3 faza repolaryzacji)
odstęp QT - wyraża czas potencjału czynnościowego mięśnia komór (depolaryzacja + repolaryzacja)
Rys.(z konspektu)
Test wysiłkowy - badanie medyczne opierające się na zależności zmieniającego się zapisu EKG pod wpływem zwiększającego się wysiłku fizycznego, który powoduje zwiększanie zapotrzebowania na tlen, co organizm kompensuje przez zwiększenie przepływu przez naczynia wieńcowe.
U osób z niewydolnością przepływu wieńcowego, powyżej pewnego wysiłku, nie może już dalej zostać pokryte to zapotrzebowanie drogą zwiększonego przepływu i rozwijają się cechy niedokrwienia mięśnia sercowego.
Wyjaśnij pochodzenie i rolę jonów Ca++ w mioplaźmie komórki serca oraz podaj czynniki wpływające na zawartość Ca++ w sercu
Gospodarka wapniowa w kardiomiocycie Stężenie wolnego Ca2+ w sarkoplazmie komórki znajdującej się w stanie spoczynku wynosi około 2 x 10-8 M. W ciągu pojedynczych milisekund po rozpoczęciu fazy 0 potencjału, jednocześnie z aktywacją kanałów wapniowych typu L, stężenie Ca2+ zaczyna szybko rosnąć i w ciągu następnych 10-20 ms osiąga wartość ~ 10-6 mol. Od szybkości zwiększania i zmniejszania stężenia Ca2+ zależy szybkość skurczu i rozkurczu. W warunkach fizjologicznych w kardiomiocytach głównym źródłem Ca2+ jest siateczka sarkoplazmatyczna. Jest to twór błoniasty składający się z dwóch morfologicznie i czynnościowo różniących się części. Część kanalikowa gęsto oplata poszczególne sarkomery. Na poziomie prążków Z część kanalikowa przechodzi w tzw. pęcherzyki końcowe, których ściany znajdują się w apozycji do ścian kanalików poprzecznych sarkolemy. Ściany pęcherzyków końcowych i ściany kanalików poprzecznych lub sarkolema ograniczają przestrzeń siateczkowo-sarkolemalna (przestrzeń SS-SL). Cały kompleks składający się z pęcherzyka końcowego siateczki, przestrzeni SS-SL oraz ściany kanalika T lub sarkolemy nosi nazwę diady (odpowiednik triady w mięśniu szkieletowym). Ściany pęcherzyków końcowych SS zawierają kanały, których aktywacja powoduje wydzielenie Ca2+ do przestrzeni SS-SL, z której dyfunduje on do sarkoplazmy. Kanały te są prawie całkowicie nieaktywne w stanie spoczynku komórki. Aktywowane są one przez wzrost stężenia Ca2+ w ich otoczeniu. Kanały wapniowe sarkolemy zlokalizowane są przede wszystkim w tych jej odcinkach, które leżą naprzeciw błony pęcherzyków końcowych SR i naprzeciw kanałów wapniowych SS po drugiej stronie szczeliny SS-SL. Kanały siateczki i sarkolemy tworzą grupy czynnościowe, w których jeden kanał SL przypada na 4 - 9 kanałów SR. W momencie aktywacji kanałów wapniowych SL, Ca2+ napływa przez nie do częściowo ograniczonej przestrzeni SS-SL. Powoduje to gwałtowne zwiększenie stężenia Ca2+ bezpośrednim otoczeniu kanałów Ca2+ siateczki i ich aktywację. Stężenie Ca2+ w przestrzeni SS-SL może dochodzić na szczycie wydzielania do 10-3 mol, tj. może ono być ok. 10 000 razy większe niż w otaczającej sarkoplazmie. Ten ogromny gradient stężeń powoduje szybką dyfuzję Ca2+ od jego źródła wzdłuż sarkomerów do miejsc wychwytu troponiny C, jak również szybkie zwiększenie jego średniego stężenia w komórce. Pociąga to za sobą szybką aktywację skurczu. Kanały wydzielające Ca2+ z SS mięśnia sercowego należą do grupy tzw. receptorów rianodynowych (RyR). Receptory rianodynowe mięśnia sercowego aktywowane są przez napływ jonów Ca2+ przez kanały Ca2+ sarkolemy. Rozkurcz jest spowodowany spadkiem stężenia Ca2+ do poziomu podprogowego dla układów kurczliwych dzięki 2 sprzężonym ze sobą procesów: Ponownego wychwytu wydzielonego Ca2+ przez ATP-azę siateczki sarkoplazmatycznej Odkomórkowego transportu na drodze wymiany Na/Ca w takiej ilości, w jakiej dyfundował on do komórki w czasie jej pobudzenia. Większość wymienników Na+/Ca2+ jest zlokalizowana, naprzeciw pęcherzyków końcowych SS i przez to eksponowane na stężenie Ca2+ wyższe niż w głównej masie sarkoplazmy, co ułatwia odkomórkowy transport Ca2+. Aktywność tej ATP-azy jest regulowana przez sprzężony z nią fosfoproteid zwany fosfolambanem. Nieufosforylowany fosfolamban wywiera na ATP-azę silny wpływ hamujący. Ca2+-ATP-aza SS jest bardzo aktywna, dzięki czemu Ca2+ jest intensywnie wychwytywany przez SS z sarkoplazmy. Ca2+ wychwytany przez kanaliki SS jest transportowany do jej pęcherzyków końcowych, gdzie jest on magazynowany częściowo w połączeniu ze specyficznym białkiem, kalsekwestryną. Aktywność tej ATP-azy jest tak duża, że wychwytuje ona cały Ca2+ napływający przez aktywowane w czasie pobudzenia kanały Ca2+ sarkolemy. Jest on przechowywany do następnego pobudzenia w pęcherzykach końcowych, a następnie wydzielany do sarkoplazmy, gdzie aktywuje skurcz. Część wydzielonego Ca2+ w ilości odpowiadającej napływowi jest usuwana do środowiska zewnątrzkomórkowego przez wymianę Na/Ca. Reszta, wraz z Ca2+, który napłynął w czasie danego pobudzenia, jest z powrotem wyłapywana przez siateczkę i znowu przetrzymywana do następnego pobudzenia. W warunkach wysokiego stężenia Ca2+ cząsteczki troponiny przyłączają po cztery jony wapnia. Wysycona jonami wapnia troponina zmienia kształt i ujawnia mostki przejściowe na powierzchni cząsteczki tropomiozyny. Pomiędzy aktyną i miozyną tworzą się mostki poprzeczne.W warunkach spoczynkowych troponina nie jest związana z jonami wapnia; dlatego też istnieje możliwość tworzenia dodatkowych mostków poprzecznych. Czynniki zwiększające stężenie wewnątrzkomórkowe Ca2+ zapewniają tworzenie się większej liczby mostków. Wzrastająca liczba mostków zwiększa siłę skurczu Kurczliwość Siła skurczu może być stopniowo zmieniana i zależy od wewnątrzkomórkowej koncentracji Ca2+. Zjawisko to jest nazywane dodatnim (efektem) inotropowym lub zwiększoną kurczliwością.
5) Co to jest objętość wyrzutowa, pojemność minutowa i wskaźnik sercowy i jakie są ich prawidłowe wartości
Objętość wyrzutowa serca (SV - stroke volume) ilość krwi wtłaczanej przez jedną z komór serca do odpowiedniego zbiornika tętniczego.
U dorosłego mężczyzny ( 70 kg, pozycja leżąca) - 75 ml.
W końcu skurczu pozostaje około 50 ml krwi stanowiącej objętość krwi zalegającej (RBV - residual blood volume) warunkującej objętość późnoskurczową komór (EVV - end-systolic ventricular volume - SV + RBV).
Frakcja wyrzutowa lewej komory SV/EVV - 75ml/75ml + 50 ml wskaźnik stanu czynnościowego lewej komory
Pojemność minutowa serca - ilość krwi tłoczonej przez jedna z komór serca w czasie jednej minuty - 5.4 L/min (90 ml/s)
Wskaźnik sercowy (cardiac index) - pojemność minutowa przeliczona na 1 m2 powierzchni ciała - 3.2 L/min/ m2 (53 ml/s/m2)
Wpływ nerwu błędnego na czynność serca
Do najważniejszych zmian czynności serca zachodzących pod wpływem stymulacji sercowych włókien nerwów błędnych lub acetylocholiny należą:
zwolnienie lub całkowite zahamowanie rytmu węzła SA i AV (ujemne działanie chronotropowe);
zmniejszenie szybkości przewodzenia, aż do jego całkowitego zniesienia w obrębie węzła AV (działanie dromotropowe ujemne);
zmniejszenie kurczliwości, głównie mięśni przedsionków (działanie inotropowe ujemne).
Nerwy błędne mają niewielki wpływ na kurczliwość mięśni komór.
Wpływ amin katecholowych na czynność serca
Katecholaminy wywołują ogólne reakcje organizmu, które przygotowują ciało do wysiłku fizycznego związanego z walką lub ucieczką. Typowe efekty działania to: podniesienie ciśnienia krwi, przyspieszenie akcji serca, a także podniesienie poziomu glukozy we krwi.
Aminy katecholowe wywierają silny wpływ na układ krążenia, a mianowicie zwiększają siłę i częstość skurczów serca (za pośrednictwem receptorów beta) oraz działają na czynność mięśni gładkich naczyń tętniczych (r. alfa i beta).
8) Od czego zależy siła skurczu serca, jakie znasz czynniki inotropowe dodatnie i ujemne
Regulacja siły skurczu mięśnia sercowego:
Istnieją dwa podstawowe mechanizmy regulacji siły skurczu (Po) mięśnia sercowego:
Przez zmianę rozkurczowej długości mięśnia.
Przez wpływ na jego kurczliwość.
Prawo Franka-Starlinga
mówi, że energia wytwarzana przez serce w czasie skurczu jest funkcją końcowo-rozkurczowego rozciągnięcia włókien mięśniowych.
Jednym z głównych czynników decydujących o sile skurczu jest początkowa długość włókien mięśniowych (ang. preload - obciążenie wstępne).
W zdrowym sercu obciążenie wstępne zależy od objętości krwi w komorach
tuż przed rozpoczęciem skurczu , która jest proporcjonalna zarówno do ciśnienia końcowo-rozkurczowego, jak i do ośrodkowego ciśnienia żylnego.
Kurczliwość
Pod pojęciem kurczliwości mięśnia sercowego rozumiemy jego podstawową zdolność do generowania siły.
Za miarę kurczliwości przyjmujemy siłę, jaką może on generować w skurczu całkowicie izometrycznym (izowolumetrycznym w sercu in situ).
Czynniki i interwencje zmieniające kurczliwość mięśnia sercowego nazywamy czynnikami czy interwencjami inotropowymi, odpowiednio dodatnimi i ujemnymi..
Stan kurczliwości mięśnia sercowego przejawia się w szybkości narastania siły i w szybkości rozkurczu.
Im większa jest kurczliwość, tym większa jest siła oraz szybkość jej narastania.
Dzięki temu następuje skrócenie czasu od początku do szczytu skurczu.
Rozkurcz trwa również krócej, co wpływa na skrócenie fazy rozkurczu izowolumetrycznego w sercu in situ.
Regulacja siły skurczu mięśnia odbywa się przez kontrolę nad ilością utworzonych wiązań aktyna-miozyna. Ilość tych wiązań zależy od ilości jonów wapniowych w sarkoplazmie.
inotropowy (gr. ís, inós `mięsień' + trópos `zwrot, kierunek') fizjol. czynniki. - związek chem., który powoduje zmianę kurczliwości serca, dodatni - przyspiesza ją, ujemny - spowalnia, np. adrenalina, inozyna (+), acetylocholina, adenozyna (-).
Antagonistyczne działanie układu autonomicznego na serce:
Układ przywspółczulny uwalnia acetylocholinę (ACh), co zmniejsza przepuszczalność dla Ca2+ i zwalnia pracę serca.
Układ współczulny uwalnia noradrenalinę (NE), co otwiera kanały wapniowe i zwiększa siłę skurczu oraz przyśpiesza pracę serca.
Charakterystyka krążenia wieńcowego
Krążenie wieńcowe
Tętnice wieńcowe;
Naczynia powierzchniowe.
Główne naczynia wieńcowe przebiegają przez nasierdzie i oddają gałęzie wnikające do mięśnia sercowego.
Odgałęzienia.
Odgałęzienia tętnic powierzchownych wnikające w mięsień serca rozdzielają się, tworząc „arkady'' zaopatrujące w krew mięsień sercowy.
W warunkach fizjologicznych tętnice wieńcowe stanowią tętnice końcowe, jednakże w przypadku zamknięcia światła wytwarza się krążenie oboczne.
Zaopatrzenie tętnic wieńcowych
Lewa:
85% krwi przepływa przez lewą tętnicę wieńcową zaopatrując krwią lewy przedsionek, przednią i większą część tylnej ściany komory lewej, część przedniej ściany komory prawej, przednie 2/3 przegrody
Prawa:
Prawa tętnica wieńcowa zaopatruje pozostałe części mięśnia sercowego.
Prawa komora
Prawy przedsionek
Tylna 1/3 przegrody międzykomorowej
Główna cz. układu przewodzącego
Tętnice wieńcowe tworzą gęste rozgałęzienia.
Drobne tętniczki są zespolone ze sobą we wszystkich warstwach mięśnia sercowego.
Liczba tętniczek oraz zespoleń rośnie przy systematycznych wysiłkach fizycznych.
Tętnice wieńcowe są bardziej podatne do rozciągania, niż tętnice mięśni szkieletowych.
Wzrost ciśnienia tętniczego wywołuje bierne rozciąganie ścian tętnic wieńcowych i zmniejszenie oporu przepływu krwi.
W tych warunkach przepływ przez tętnice wieńcowe może wzrosnąć w stopniu 4 do 5 razy większym niż w tętnicach zaopatrujących mięśnie szkieletowe.
Liczba naczyń włosowatych w mięśniu sercowym jest 3 do 4 razy większa, niż w mięśniu szkieletowym.
Na każde włókno mięśniowe przypada jedno naczynie włosowate.
Żyły wieńcowe
Krew żylna odpływa z serca:
Układ naczyń żylnych powierzchownych
Układ naczyń żylnych głębokich
Żyłami Tebezjusza
Żyły
Żyły zbierające krew z dorzecza lewej tętnicy wieńcowej uchodzą do żyły wieńcowej, a ta przechodzi ku tyłowi w zatokę wieńcową.
Żyły Tebezjusza i żyły najmniejsze serca doprowadzają krew żylną do lewej komory, tworząc przeciek.
Żyły zbierające krew z dorzecza prawej tętnicy wieńcowej to liczne przednie żyły wieńcowe.
Na połączenie pomiędzy naczyniami wieńcowymi a światłem komory w prawej komorze przypada część przepływu większa niż w lewej
Przepływ wieńcowy krwi.
Ciągły dopływ krwi do serca stanowi podstawowy warunek utrzymania wymaganej podaży O2 i substancji odżywczych.
Typowy przepływ wieńcowy stanowi około 5% spoczynkowej pojemności minutowej serca (250 ml/min) lub w przybliżeniu 60-80 ml krwi/100 g tkanki/min.
Zadanie krążenia wieńcowego:
Zaopatrzenie serca w krew proporcjonalnie do jego zapotrzebowania na O2 zależne od:
Stopnia ekstrakcji tlenem
Ilości krwi dostarczanej w jednostce czasu
Przepływ wieńcowy zależy od :
częstotliwości skurczów serca
ciśnienia końcowo - rozkurczowego w aorcie
ciśnienia końcowo - rozkurczowego w komorze
oporu obwodowego w ogólnym krążeniu, czyli afterload
Komponenta naczyniowa
Autoregulacja przepływu wieńcowego
Regulacja metaboliczna - przekrwienie czynne
Rola śródbłonka w regulacji szerokości naczyń
11) Czynniki regulujące napięcie mięśniówki oskrzeli:
Istotną rolę w regulacji napięcia mięśni gładkich oskrzeli odgrywają włókna autonomiczne nieadrenergiczne i niecholinergiczne NANC gałązek eferentnych nerwu błędnego:
Włókna hamujące NANC; rozluźniają napięcie mięśni gładkich i rozszerzają oskrzela.
Działają one za pośrednictwem tlenku azotu (NO), rozszerzającego oskrzela w pierwszej fazie rozkurczu.
W drugiej, późniejszej, ale długiej fazie rozkurczu rozszerzają one oskrzela za pośrednictwem neuropeptydów: wazoaktywnego peptydu jelitowego (VIP) i peptydów przysadkowych aktywujących cyklazę adenylową (PACAP).
Włókna pobudzające NANC, kurczące mięśnie gładkie oskrzeli przez neurokininy uwalniane w zakończeniach, głównie substancję P działającą na receptor NK1 oraz neurokininę A (NKA) działającą na receptor NK2.
Zarówno hamujące, jak i pobudzające transmittery NANC wydzielane są na zasadzie kotransmisji wraz z acetylocholiną i transmitterami zawracających kolaterali
Trzewnoczuciowych włókien peptydergicznych nerwu błędnego uwalniających m.in. cCGRP.
12 Zasady wymiany dwutlenku węgla i tlenu w płucach i tkankach, formy ich transportu w płynach ustrojowych, rola fizjologiczna i zawartość w organizmie.
Wymiana gazowa w płucach: Powietrze jest mieszaniną gazów. Najważniejszymi składnikami są: azot ok. 78%, tlen ok. 21%, dwutlenek węgla ok. 0,03% i inne gazy. Z powietrza wciągniętego do wnętrza pęcherzyków płucnych tlen przechodzi do naczyń włosowatych, a dwutlenek węgla, przetransportowany przez krew, w odwrotnym kierunku, do wnętrza pęcherzyka płucnego, a następnie do oskrzeli, tchawicy, krtani - na zewnątrz.
Jest to zjawisko fizyczne oparte na zasadach dyfuzji. Każdy gaz przechodzi z miejsca, gdzie znajduje się w wyższym stężeniu, do miejsca o niższym stężeniu. Tlen pobrany w pęcherzykach płucnych łączy się z hemoglobiną. Proces ten nazywamy zewnętrzną wymianą gazową.
Mechanizm wentylacji płuc:
Wdech:
mięśnie międzyżebrowe kurczą się,
żebra unoszą się do góry,
zwiększa się objętość klatki piersiowej,
przepona spłaszcza się.
Wydech:
mięśnie międzyżebrowe rozluźniają się,
żebra wracają do pierwotnego położenia,
zmniejsza się objętość klatki piersiowej.
Ilość wydychanego powietrza zależy od:
tempa oddychania,
głębokości oddychania.
Mechanizm oddychania zachodzi niezależnie od naszej woli. Kieruje nim układ nerwowy, którego ośrodki znajdują się w rdzeniu przedłużonym.
Wymiana gazowa w tkankach:
Tlen pobrany w pęcherzykach płucnych transportowany przez krew, dociera do tkanek. Tam odbywa się wymianagazowa wewnętrzna, tzn. tlen przenika do komórek, a z komórek pobierany jest dwutlenek węgla, który z krwią wędrować będzie do płuc. Wędrówka gazów w tkankach odbywa się również na zasadzie dyfuzji
Główną funkcją krwi jest transport gazów oddechowych: tlenu i dwutlenku węgla. Oba gazy przenikają przez błony komórkowe i ściany naczyń krwionośnych wskutek gradientu ciśnienia parcjalnego.
Tlen przechodzi do krwi z płuc. Tam wiązany jest w postaci cząsteczkowej przez hemoglobinę - czerwony barwnik erytrocytów. Jedna cząsteczka hemoglobiny może związać cztery cząsteczki tlenu - powstaje kompleks zwany oksyhemoglobiną (hemoglobina utlenowana) Hb(O2)4. Gazem o powinowactwie do hemoglobiny wyższym niż tlen jest tlenek węgla (czad, CO). Wypiera on tlen z oksyhemoglobiny i łączy się z nią, tworząc karoksyhemoglobinę, co jest niebezpieczne dla organizmu - powoduje zablokowanie dopływu tlenu do tkanek i śmierć w wyniku uduszenia.
Dwutlenek węgla przechodzi do krwi z komórek, gdzie powstaje jako produkt metabolizmu. Tylko 20% tego gazu transportowane jest przez hemoglobinę - są to luźne połączenia z grupami -NH2 (karbaminohemoglobina). Pozostała część (80%) transportowana jest w osoczu w postaci jonów wodorowęglanowych HCO3-.
13) Regulacja dowolna oddychania, jej mechanizmy i ograniczenia
MECHANIZMY REGULACJI ODDYCHANIA NA POZIOMIE O.U.N.
Oddychanie regulują dwa niezależne mechanizmy.
Jeden z nich jest odpowiedzialny za kontrolę dowolną, drugi za automatyczną.
Układ związany z oddychaniem dowolnym jest zlokalizowany jest w korze mózgowej i wysyła impulsy do motoneuronów oddechowych poprzez drogi korowo-rdzeniowe.
Mięśnie oddechowe są typowymi mięśniami szkieletowymi, pobudzanymi przez o.u.n. za pośrednictwem odpowiednich nerwów ruchowych.
Unerwienie. Skurcz mięśni oddechowych jest wynikiem aktywności unerwiających je nerwów somatycznych.
Układ związany z kontrolą automatyczną znajduje się w moście i w rdzeniu przedłużonym.
Wypustki eferentne tego układu, biegnące do motoneuronów oddechowych, znajdują się w istocie białej rdzenia kręgowego pomiędzy bocznymi i przednimi drogami korowo-rdzeniowymi.
Włókna związane ze wdechem konwergują na motoneuronach przeponowych, rozmieszczonych od C3 - C5 , oraz na motoneuronach unerwiających m. międzyżebrowe zewnętrzne, zlokalizowanych w rogach przednich w obrębie segmentów piersiowych r. kręgowego.
Włókna odpowiedzialne za wydech konwergują przede wszystkim na motoneuronach w odcinku piersiowym r. kręgowego, unerwiających m. międzyżebrowe wewnętrzne.
14) Rodzaje i rola receptorów płucnych
Receptory płucne dzieli się na 4 rodzaje:
Wolno adaptujące mechanoreceptory (SAR - Slow adapting receptors) znajdujące się w tchawicy i oskrzelach, wrażliwe na rozciąganie płuc (receptory inflacyjne - rozciągowe) w czasie wdechu.
Receptory szybko adaptujące się płuc (RAR - rapid adapting receptors), zwane także podnabłonkowymi, wrażliwe na bodźce chemiczne w drogach oddechowych, czyli receptory typu I (Irritant receptors)
Receptory okołokapilarne (J-Juxtacapillary receptors) zlokalizowane w przegrodach pomiędzy kapilarami pęcherzykowymi a pneumocytami.
Receptory oskrzelowe włókien aferentnych typu C, obecne w całym drzewie oskrzelowym, wrażliwe na autokoidy płucne (np. histamina, leukotrieny, tachykininy i kapsaicyna).
Mechanoreceptory SAR (inflacyjne) Należą do receptorów wolnoadaptujących, czyli utrzymują wysoką częstość wyładowań mimo dłuższego rozciągania płuc. Wynikiem pobudzenia tych receptorów podczas wdechu jest odruchowe zahamowanie tzn. skrócenie i spłycenie wdechu oraz przyspieszenie rytmu oddechowego. Odruch z udziałem tych receptorów, zwany odruchem Heringa-Breuera lub inflacyjnym, jest najważniejszym czynnikiem regulacji oddychania przez nerwy obwodowe i po jego wyeliminowaniu przez przecięcie nerwów błędnych wdechy stają się głębsze i dłuższe. Wzrasta objętość oddechowa i rytm oddechowy staje się wolniejszy. Główną rolą odruchu Heringa-Breuera jest ujemne sprzężenie zwrotne ograniczające czas trwania wdechu przez pobudzenie neuronów P w grupie neuronów grzbietowych DRG-NTS wyłączających wdech. Poza odruchem Heringa-Breuera typu inflacyjnego istnieje także deflacyjny odruch Heringa-Breuera (pobudzająco-wdechowy), inicjowany przez spadek aktywności tych samych receptorów SAR, związanych z odruchem inflacyjnym lub przez pobudzenie innych receptorów (deflacyjnych) na skutek zapadania się płuc./ Informacje z tych receptorów docierają poprzez nerwy błędne do ośrodków oddechowych pnia mózgu, prowadząc do zmiany wydechu we wdech. Jego rola u dorosłych (gdy objętość oddechowa przekroczy 1l) sprowadza się raczej do dostosowania czasu trwania i głębokości wdechów do właściwości mechanicznych (podatność) płuc i klatki piersiowej.
Receptory szybko adaptujące RAR lub typu I Pobudzenie tych receptorów wywołuje odruch pobudzający aktywność oddechową z pogłębieniem i przyspieszeniem ruchów oddechowych, czyli hiperwentylację. Ponadto pobudzenie receptorów RAR wywołuje odruch kaszlu i skurcz oskrzeli. Ponieważ te receptory ulegają pobudzeniu przez histaminę i inne autokoidy uwalniane np. w astmie, przypisuje się im także rolę w mechaniźmie skórczu oskrzeli. U drosłych rola fizjologiczna receptorów RAR polega także na odruchowym przeciwdziałaniu spadkowi podatności płuc i gorszemu upowietrznianiu pęcherzyków płucnych.
Receptory typu J to wolne zakończenia w ścianie pęcherzyków płucnych w pobliżu naczyń kapilarnych. Efektem odruchów z receptorów J jest krótkotrwały bezdech, a następnie płytkie i częste ruchy oddechowe. Towarzyszy im odruchowe zwężenie oskrzeli i skurcz mięśni zamykających głośnię oraz pobudzenie nerwów błędnych, zwolnienie akcji serca, zmniejszenie napięcia naczynioruchowych nerwów adrenergicznych i rozszerzenie naczyń krwionośnych. Odruchy z receptorów J mają charakter obronny, gdyż ostatecznie zmniejszają napływ czynników szkodliwych (zamknięcie głośni i skurcz oskrzeli, spłycenie oddechów) do płuc. Te receptory mają być także odpowiedzialne za uczucie duszności, czyli „krótki” oddech, występujący w chorobach płuc.
Pewne znaczenie w mechanizmach odruchowych przypisuje się bardzo licznym cienkim włóknom C w płucach. Są one odpowiedzialne za odruchy wywołane autokoidami i kapsaicyną oraz biorą udział w patogenezie astmy, zapalenia i obrzęku płuc. Reakcjom odruchowym, w których pośredniczą włókna C towarzyszy bezdech a potem płytkie i częste ruchy oddechowe, bradykardia i spadek ciśnienia krwi. Informacje przekazywane włóknami C są subiektywnie odczuwane jak uczucie bólu, ucisku i palenia, które towarzyszą działaniu substancji drażniących na płuca.
Chemoreceptory obwodowe i centralne, mechanizmy ich aktywacji i ich rola w regulacji oddychania
Kontrola chemiczna
Centralna
Chemoreceptory ośrodkowe są neuronami umiejscowionymi tuż pod brzuszną powierzchnią opuszki mózgu.
Neurony te zmieniają swoją aktywność bioelektryczną w zależności od stężenia H+ w płynie mózgowo-rdzeniowym.
Około 85% spoczynkowego napędu oddechowego pochodzenia chemicznego jest uwarunkowane pobudzającym wpływem CO2 (H+) na chemoreceptory ośrodkowe.
Przy wzroście PCO2 i obniżeniu pH dochodzi do zwiększenia wentylacji przez pogłębienie oddechów i zwiększenie częstości oddychania w wyniku pobudzenia ośrodka oddechowego za pośrednictwem strefy chemowrażliwej mózgu (CSA, chemosensi-tive area).
Strefa ta znajduje się na brzusznej powierzchni opuszki i dzieli się na strefę do-głowową (strefa R) i strefę doogonową (strefa C).
Strefy te rozciągają się do mostu, z boku do korzeni nerwów VII-X, przyśrodkowo do szlaków piramidowych, a doogonowo do korzeni nerwu XII.
Neurony obu stref przekazują pobudzenia chemiczne z chemowrażliwych receptorów do opuszkowego ośrodka oddechowego.
Obwodowa
Istnieją specjalne receptory poza ośrodkowym układem nerwowym, które reagują na zmiany ciśnienia parcjalnego O2 i CO2 oraz wzrost stężenia jonów we krwi tętniczej.
Chemoreceptory obwodowe
Ich pobudzenie, występujące przy obniżonym ciśnieniu parcjalnego O2 (hipoksemia), niskim pH (kwasica) i podwyższonym ciśnieniu parcjalnym CO2 (hiperkapnia) we krwi tętniczej, przekazywane jest do ośrodka oddechowego włóknami czuciowymi nerwu IX i X.
Chemoreceptory zgrupowane są w obrębie kłębków umiejscowionych symetrycznie w pobliżu rozwidlenia tętnicy szyjnej wspólnej (kłębki szyjne) i w łuku aorty w liczbie 3-4 (kłębki aortalne).
Są one zbudowane z tzw. kłębuszków, zaopatrzonych w tętniczki, które dzielą się na liczne naczynia włosowate i sinusoidalne, otoczone zewsząd komórkami kłębkowymi typu nabłonkowego (komórki typu I).
Komórki typu I wyposażone są w wypustki stykające się ze ścianami pobliskich naczyń kapilarnych i z sąsiednimi komórkami typu II.
Komórki typu II w kłębkach pełnią funkcje podporowe o charakterze podobnym do funkcji komórek glejowych.
Ziarnistości w komórkach typu I gromadzą aminy katecholowe, głównie dopaminę, noradrenalinę i serotoninę oraz substancję P, VIP i enkefaliny.
Komórki kłębków szyjnych wykazuje najwyższe zużycie O2 (ok. 9 ml/100 g tkanki/min).
Czerpią go z postaci rozpuszczonej fizycznie w osoczu krwi, która przepływa tu w dużych ilościach (aż ok. 2000 ml/100 g tkanki/min, czyli 40-krotnie więcej niż w mózgu).
5
Wyszukiwarka
Podobne podstrony:
Kopia Fizjologia - odpowiedzi 1-15, PIELĘGNIARSTWO ROK 1 LICJENCJAT, FIZJOLOGIA
Kolokwium II - Fizjologia 16-30, PIELĘGNIARSTWO ROK 1 LICJENCJAT, FIZJOLOGIA
Recertory jonotropowe i metabroponoe, PIELĘGNIARSTWO ROK 1 LICJENCJAT, FIZJOLOGIA
FIZJOLOGIA UKĹADU NERWOWEGO 5, PIELĘGNIARSTWO ROK 1 LICJENCJAT, FIZJOLOGIA
Kolokwium II fizjologia pytania
petagogika wykład, PIELĘGNIARSTWO ROK 1 LICJENCJAT
Temat - filozofia.pop, PIELĘGNIARSTWO ROK 1 LICJENCJAT, praca poprawiona
WOK Zagadnienia na kolokwium II semestr, Kulturoznawstwo UG, rok I
Układ oddechowy - kolokwium 2, II rok, fizjologia
Fizjologia - układ oddechowy - odpowiedzi do pytań z kolokwium, II rok, II rok CM UMK, Fizjologia, F
fizjologia roslin - kolokwium, biotechnologia 2 sem rok2, pobrane z góry DS 7, z góry, Rok II, Fizjo
układ stawowy i układ mięśniowy, Pielęgniarstwo rok I i inne, Anatomia i Fizjologia
dla ciekawych wtorku, Rok III, Rok II, Fizjologia i żywienie zwierząt, Fizjologia i żywienie zwierzą
Fizjologia kolokwium II
Fizjologia3, biotechnologia 2 sem rok2, pobrane z góry DS 7, z góry, Rok II, Fizjologia roślin
fizjologia kolos 2, BIOLOGIA UJ LATA I-III, ROK III, semestr II, Fizjologia roślin
Fizjologia2, z góry, Rok II, Fizjologia roślin
więcej podobnych podstron