FIZJOLOGIA II
1. CO TO JEST FITOCHROM?
FAZA CIEMNA FOTOSYNTEZY PRZEBIEG I ZNACZENIE.
Ca, Mg, N, ZNACZENIE W FIZJOLOGII.
CO TO SĄ GLIOKSYSOMY ZNACZENIE W METABOLIŹMIE.
MECHANIZM KWITNIENIA.
MECHANIZM TRANSPORTU W KSYLEMIE I FLOEMIE
CYKL HATCHA I SLACKA ( C4)
SUBSTANCJE PRZEWODZONE W KSYLEMIE I FLOEMIE I MECHANIZM ICH TRANSPORTU.
OD JAKICH CZYNNIKÓW ZALEŻY WRAŻLIWOŚĆ ROŚLIN NA NIEKORZYSTNE WARUNKI ŚRODOWISKA.
10.FOTOODDYCHANIE.
11.OPIS CYKLU KALVINA BENSONA W FOTOSYNTEZIE.
12.PODZIAŁ BIAŁEK ENZYMATYCZNYCH.
GIBERELINA I AUKSYNA ICH ZNACZENIE PRZY RÓŻNICOWANIU WZROSTU I SŁOJÓW.
14. ZNACZENIE Fe, K, Mg W METABOLIŹMIE.
15. CO TO JEST GLIKOLIZA I JAKIE JEST JEJ ZNACZENIE W PROCESACH KATABOLICZNYCH ROŚLIN ?
16. OMÓW REAKCJE FOTOPERIODYCZNE U ROŚLIN DRZEWIASTYCH.
17. MECHANIZM TRANSPORTU BIERNEGO I CZYNNEGO
18.CO OZNACZA TERMIN STRES W FIZJOLOGII ROŚLIN.
19.CYKL KREBSA - PRZEBIEG I ZNACZENIE.
20.INHIBITORY WZROSTU.
21.CO TO JEST FIZJOLOGIA ?
22. ZNACZENIE TRANSPIRACJI.
23. ZNACZENIE PROCESÓW REDOKS W PROCESIE METABOLIZMU.
24. CO TO JEST POTENCJAŁ WODY I JAKIE JEST JEGO ZNACZENIE ?
25. OD CZEGO ZALEŻY SZYBKOŚĆ REAKCJI ENZYMATYCZNEJ ?
26. MIEJSCE POWSTAWANIA I PROCES PRZEMIESZCZANIA SIĘ
STYMULATORÓW WZROSTU.
27. TRANSPORT ELEKTRONÓW I ZNACZENIE W METABOLIŹMIE
WPŁYW FITOCHROMÓW NA ODPORNOŚĆ ROŚLIN , NA NIEKORZYSTNY WPŁYW ŚRODOWISKA.
29.ROLA CZYNNIKÓW HORMONALNYCH W PROCESACH REGENERACJI ROŚLIN DRZEWIASTYCH.
30. ZNACZENIE GIBERELINY W PROCESIE WZROSTU I RÓŻNICOWANIA DRZEW.
31.ZNACZENIE FOTOINDUKCJI I TERMOINDUKCJI W PROCESIE WZROSTU I ROZWOJU ROŚLIN.
32.ZNACZENIE N I Mg W METABOLIŹMIE
33.OGÓLNA CHARAKTERYSTYKA I ZNACZENIE PROCESÓW KATABOLICZNYCH I ANABOLICZNYCH
34. MECHANIZM TRANSPORTU WE FLOEMIE
35. ZNACZENIE P , WYSTĘPOWANIE.
36.MIEJSCE SYNTEZY , DROGI I MECHANIZMY TRANSPORTU AUKSYNY.
37. CZYNNIKI WARUNKUJĄCE INTENSYWNOŚĆ FOTOSYNTEZY U DRZEW LEŚNYCH.
1.CO TO JEST FITOCHROM?
Fitochrom jest to barwnik, który uczestniczy w procesie związanym z pochłanianiem widma słonecznego w zakresie światła czerwonego i dalekiej czerwieni.
Obecność fitochromu stwierdzono u wszystkich roślin. Barwnik ten funkcjonuje w dwóch formach molekularnych : Pr - wykazuje maksimum absorbcji w zakresie światła czerwonego - 660 nm , aPfr - pochłania światło dalekiej czewieni - 730 nm.(Może dochodzić do przekształcenia jednej formy w drugą). Izolowany fitochrom jest rozpuszczalną w wodzie chromoproteiną o barwie błękitnej. Fitochrom jest zbudowany z polipeptydu połączonego z grupą chromoforową. Wyróżniono dwie fizjologiczne populacje fitochromu : labilną (PI) i stabilną (PII). Fitochrom labilny oznacza się jako fitochrom A , a stabilny jako fitochrom B. Przyjmuje się, że tylko Pfr jest aktywną formą fitochromu . Pfr i Pr absorbują światło czerwone ale daleką czerwień już tylko Pfr. Pochłonięciu światła przez Pr towarzyszy uruchomienie łańcucha przemian metabolicznych.
Przypuszcza się,że fitochrom labilny i stabilny działają niezależnie od siebie lub tez współdziałają w kontroli wzrostu i rozwoju roślin. Fitochrom kontroluje także deetiolację (przejście od heterotroficznej do autotroficznej formy odżywiania roślin).
Procesy w których pośredniczy fitochrom to :
stymulacja kiełkowania
hamowanie wydłużania się łodygi
stymulacja wzrostu powierzchni liści
stymulacja kwitnienia
FAZA CIEMNA FOTOSYNTEZY PRZEBIEG I ZNACZENIE.
Stroma to miejsce fazy ciemnej fotosyntezy w której ATP i NADPH wytworzone w fazie świetlnej są używane do redukcji CO2 i wbudowania go w związki organiczne. Oprócz enzymów zaangażowanych w asymilację CO2 ( cykl Calvina Bensona ) w stromie znajdują się DNA ( genom) chloroplastowy oraz plastytowy aparat biosyntezy białka. Reakcje ciemnej fazy zachodzą w stromie gdzie znajdują się wszystkie enzymy uczestniczące w tym procesie. W stromie zachodzi proces karboksylacji katalizowany przez karboksylazę 1,5-bifosforybulozy. Siła asmilacyjna redukuje grupę karboksylową wytworzoną po związaniu CO2 do grupy aldehydowej czyli do poziomu cukrów. Przemiany fazy ciemnej mają charakter cykliczny i noszą nazwę cyklu Calvina. W cyklu Calvina rozróżnia się 3 fazy :
karboksylacja - przyłączenie CO2 do odpowiednich związków organicznych. Karboksylacja związana z aktywnością 1,5 bifosforybulozy i utworzeniem 2 cząsteczek kwasu 3-fosfoglicerynowego (PGA). Reakcja ta przebiega ze spadkiem energii swobodnej co powoduje jej nieodwracalność nawet gdy stężenie CO2 jest małe.
PGA - trwały produkt wiązania CO2 ulega przemianom w związki pośrednie służące do syntezy aminokwasów, białek, tłuszczów. Zawiera grupę COOH.
Redukcja - do zredukowania związków organicznych potrzeba wodoru i energii. Czynników tych dostarcza NADPH ( wodoru) ATP ( energii). Faza redukcji polega na wytworzeniu fosfotriozy (aldehyd 3 fosfoglicerynowy). Uczestniczą takie enzymy jak kinaza fosfoglicerynianowa i dehydrogenaza aldehydu 3- fosfoglicerynowego.
Regeneracja- odtwarza się związek wiążący CO2 ( 1,5-bifosforybulozy). Wymaga to udziału enzymów: aldolazy, transketolazy, fosfatazy, izomerazy, epimerazy i kinazy.
Ca, Mg, N, ZNACZENIE W FIZJOLOGII.
Ca- występuje w glebie jako rozpuszczalny, wymienny ,niewymienny. Przyczynia się do tworzenia struktury gruzełkowatej. Czynnik żyzności gleby. Pobierany w postaci jonu przenoszony do wierzchołków roślin - liści. Nadmiar szkodliwy. W roślinie występuje mitochondriach, w pektynianach wapnia, ścianach komórkowych, blaszce środkowej. Inkrustuje powierzchnię ścian. Wpływa na strukturę komórki, jest antagonistą potasu. Zły stosunek Ca do K zaburza funkcjonowanie. Wapń utrzymuje strukturę membran cytoplazmatycznych. Wpływa na metabolizm. Utrzymuje biokoloidy komórkowe, enzymy, struktury membran plazmatycznych, udział w organizacji fizjologicznej komórki. Wpływa na odporność i wzrost ( odporność na warunki klimatyczne). Zwiększa wytrzymałość ścian komórkowych.
Mg- występuje w glebie jako rozpuszczalny, wymienny i niewymienny. W roślinie w chlroplastach, rybosomach, związany z koloidami struktur komórkowych. W związkach chemicznych takich jak: chlorofil, pektyniany, szczawiany, fityna. Związki chelatowe- połączenia metali z dużą cząsteczką związku organicznego. Struktury te wzmacniają i stabilizują cząsteczkę. Magnez aktywizuje układ enzymatyczny. Bierze udział w glikolizie, cyklu kwasu cytrynowego, syntezie materiałów budulcowych, zapasowych, przemianie związków azotowych, wpływa na stan biokoloidów komórek, utrzymuje strukturę rybosomów.
N- występuje w związkach organicznych - mocznik, białko, aminokwasy, amidy, aminy, kwasy nukleinowe, próchnica. W związkach nieorganicznych- sole amonowe i azotanowe. Jako azot cząsteczkowy dostępny dla mikroorganizmów oraz roślin żyjących z nimi w symbiozie. W roślinach białka enzymatyczne, strukturalne, zapasowe, pierścienie heterocykliczne zbudowane z węgla i azotu, zasady amoniowe, zasady azotowe ( adenina, guanina, cytozyna, tymina).
Niebiałkowe związki azotu :nukleotydy, porfiryny, fosfolipidy, alkaloidy, substancje cyjanogenne, olejki gorczynne.
CO TO SĄ GLIOKSYSOMY ZNACZENIE W METABOLIŹMIE.
Glioksysomy - mikrociała charakterystyczne tylko dla roślin, nie wiadomo w jaki sposób powstają. Mogą przekształcać się w peroksysomy. Glioksysomy przekształcają się w peroksysomy przez zanik enzymów cyklu glioksalanowego i wzrost aktywności enzymów szlaku glikolanowego. Obecność glioksysomów stwierdza się wyłącznie w liścieniach oraz komórkach bielma oraz warstwie aleuronowej nasion wyspecjalizowanych w gromadzeniu tłuszczów jako materiału zapasowego. Zawarte w glioksysomach umożliwiają przekształcenie lipidów w cukry wykorzystywane podczas kiełkowania nasion przez rozwijający się zarodek.
Peroksysomy : srednica 0,1-1,5 um otoczone pojedyńczą błoną we wnętrzu obserwuje się amorficzne i włókienkowate wkręty tzw. nukleoidy.
Glioksysomy zawierają enzymy cyklu glioksalowego umożliwiają szybki rozkład kwasów tłuszczowych i zamianę na cukier. Proces ten odbywa się przy udziale mitochondriów i cytoplazmy podstawowej.
CO TO SĄ GLIOKSYSOMY ZNACZENIE W METABOLIŹMIE -druga wersja
Glioksysomy są to kuliste organelle występujące w dużych ilościach w komórkach nasion roślin oleistych. Zawarte w tych stukturach enzymy przekształcają nagromadzone w komórkach tłuszczowce zapasowe w cukry, zużywane następnie w czasie kiełkowania przez rosnący kiełek. W glioksysomach zachodzą przemiany cyklu glioksalowego, stanowiącego jeden z kilku możliwych sposobów oddechowych występujących u roślin. Cykl ten odbywa się w glioksysomach przy ścisłej współpracy z mitochondriami. W kiełkujących nasionach część rozłożonego tłuszczu stanowi substrat oddechowy służący do produkcji energii. Jest on silniej zredukowany niż cukrowce, to też utlenienie grama tłuszczu dostarcza około 40 kJ., podczas gdy utlenienie grama skrobi tylko około 16 kJ. Rozłożone tłuszcze dostarczają szkieletów węglowych stanowiących materiał do syntezy cukrowców niezbędnych w procesach wzrostu i różnicowania rozwijającej się siewki
MECHANIZM KWITNIENIA.
Po zakończeniu okresu młodocianego w którym następuje różnicowanie wegetatywne roślina wchodzi w okres rozwoju generatywnego. Czas trwania czasu juwenilnego jest właściwością genotypową. Roślina po okresie juwenilnym wchodzi w okres dojrzewania, okres kwitnienia. Warunki przejścia do stanu generatywnego.
czynniki zewnętrzne (temp. i długość dnia oraz nocy)
czynniki wewnętrzne ( wiek , rozmiar)
Jest wiele hipotez wyjaśniających dlaczego roślina zakwita. Gotowość do kwitnienia to uzyskanie przez roślinę zdolności do tworzenia poza wierzchołkiem wzrostu tzw. impulsów kwitnienia, które przemieszczając się powodują jej przejście do fazy generatywnej. Występują dwie zależności.
wernalizacja- czyli pobudzanie pod wpływem niskich temperatur na zakwitanie
termiczna indukcja kwitnienia. Wśród zielnych wyróżnia się jednoroczne ( rozwój na wiosnę, kwitnienie latem ), dwuletnie ( pierwszy rok powstają organy wegetatywne drugi rok generatywne). Zjawisko indukcji działania obniżonej temperatury na kwitnienie to wernalizacja. Temperatury indukujące to 0-10oC. Długość trwania obniżonej temperatury potrzebnej do zakwitania jest różna dla różnych roślin. Od kilkunastu do kilkudziesięciu dni. Indukcja występuje w komórce zdolnej w komórce zdolnej do podziału. Wernalizacji ulegają różne organy. Miejscempercepcji jest wierzchołek wzrostu.
fotoperiodyzm- czyli wpływ trwania dnia i nocy na zakwitanie.
Wyróżnia się następujące grupy fotoperiodyczne.
- RDK (rośliny dnia krótkiego). Kwitną gdy w dziennym fotoperiodzie jest przewaga fazy ciemnej ponad 12 godzin ciemności. ( 1 roczne kwitną jesienią)
- RDD ( rośliny dnia długiego) . Gdy w dziennym fotoperiodzie przewaga fazy jasnej ( ponad 12 godzin światła) -kwitną w lecie.
-RDKD oraz RDDK - ich inicjacja generatywna zależy od ekspozycji na dzień krótki i długi w odpowiedniej kolejności
-RN ( rośliny neutralne) - nie wrażliwe na długość dnia sygnałem dla nich jest wielkość albo wiek.
W liściach tworzą się induktory kwitnienia które przemieszczają się do wierzchołków inicjując w nich procesy generatywne ( rola czynników pokarmowych ; rola florigenu; giberyliny; cytokininy; auksyny; etylenu; kwasu abscysynowego)
MECHANIZM KWITNIENIA- druga wersja
Roślina może zakwitnąć po zakończeniu okresu juwenilnego. Merystem wierzchołkowy przestaje tworzyć liście międzywęźla wydłużają się i zaczynają się wytwarzać zawiązki kwiatowe. Wytworzenie zawiązków kwiatowych zależy od czynników wewnętrznych (wiek i odpowiedni rozmiar rośliny) i zewnętrznych (temperatura, długość dnia i nocy).w związku z tym wyróżniamy dwa procesy: wernalizację i fotoperiodyzm. Wernalizacja (termoindukcja )- pobudzający wpływ niskich temperatur na zakwitanie. Rośliny zielne można podzielić na jednoroczne i dwuletnie. Rośliny jednoroczne zaczynają rozwój na wiosnę, w lecie zawiązują kwiaty i wydają nasiona. Rośliny dwuletnie w pierwszym roku wytwarzają jedynie organy wegetatywne. Rośliny te mogą wytworzyć zawiązki kwiatowe jedynie po przejściu zimy. Na drugi rok przechodzą do stadium generatywnego. Bez okresu przechłodzenia rośliny te nie zakwitają lub zakwitają ze znacznym opóźnieniem.
Miejscem percepcji czynnika chłodu w roślinie jest merystem wierzchołkowy, a w nasionach części embrionalne. Efekt jaryzacji może być utrwalony, efekt wernalizacji może zostać przeniesiony drogą szczepienia. Hormon odpowiedzialny za kwitnienie to wernalinaniektóre rośliny rozetowe zakwitają w wyniku potraktowania gibereliną. Zastępuje to w pewnych przypadkach okres przechłodzenia w okresie wernalizacji.
Fotoperiodyzm jest reakcją rośliny na czas trwania i periodyczne następstwo podczas okresów światła i ciemności. Reakcja roślin przejawia się w postaci zakwitania a także wywarzana bulw, krzewienia, stanu spoczynku, opadania liści. Niektóre rośliny mogą zakwitać tylko w warunkach dnia długiego (14 h - lato) inne tylko w warunkach dnia krótkiego (10 h - wczesna wiosna, jesień).
Liczy się okres nieprzerwanej ciemności (reagują na światło białe i czerwone przy świetle o długości fali 730 (daleka czerwień) rośliny nie reagują na przerwanie ciemności). Liczy się ostatni błysk światła.
Hipotezy:
1.H. fitochromu - fitochrom działa bezpośrednio, zmienia przepuszczalność membran komórkowych. Zmiany przepuszczalności mają spowodować zmiany w roślinie.
2. H. Mohra - efekt fitochromu nie ogranicza się jedynie do przepuszczalności membran komórkowych, ale również do uaktywnienia niektórych genów, stymuluje syntezę określonych enzymów.
3. H. Florigenu - efekt indukcji fotoperiodycznej może być przekazywany z rośliny w drodze szczepienia. Miejscami percepcji bodźca fotoperiodycznego są rozwinięte liście.
W warunkach dnia długiego tworzą się cewki przypominające drewno wczesne, a w warunkach dnia krótkiego przypominające drewno późne.
MECHANIZM TRANSPORTU W KSYLEMIE I FLOEMIE
Naczynia ( ksylem) - transport wody i składników mineralnych. Do transportu służą naczynia i cewki oraz rurki sitowe ( do dalekiego transportu).
W ksylemie następuje przemieszczanie pomiędzy organami rośliny. Woda w ksylemie przemieszcza się do wyżej położonych części roślin. Słupy wody podciągane są do góry dzięki ssącemu działaniu parowania tworząc tzw. prąd transpiracyjny. Szybkość transportu produktów zależy od intensywności transpiracji. Intensywność transpiracji nie ma bezpośredniego wpływu na ilość substancji przetransportowanych do górnej części rośliny.
a)Parcie korzeniowe jest jednym z elementów wpływających na transpirację.
b)Transpiracja oraz teoria kohezyjno - transpiracyjna.
Siły kohezyjne powodują wzajemne przyciąganie się cząsteczek wody. Utrzymują słup pod napięciem. O szybkości przepływu decyduje powierzchnia systemu przewodzącego.
Floem- w skład floemu wchodzą 3 typy komórek sitowe, towarzyszące, miękiszowe, wzmacniające.
Floem jest podstawowym systemem przewodzącym związki organiczne na duże odległości. Transport produktów fotosyntezy składa się z 3 współzależnych od siebie etapów. Eksport asymilatów z chloroplastów do cytoplazmy ,a następnie z komórek miękiszowych liści do tkanek przewodzących to jest załadunek floemu. Następnie jest transport w samym floemie, po przemieszczeniu transportowanych związków z floemu do komórek organów - rozładunek floemu. Przewodzenie w floemie uwarunkowane jest dopływem energii metabolicznej.
Mechanizm transportu w ksylemie i floemie.- wersja druga.
Szybkość przewodzenia i ilość przewodzonych substancji. Daleki transport wody i jonów zachodzi w naczyniach i cewkach. Woda tworzy w nich nieprzerwane włosowate słupy, sięgające od korzeni aż do liści. Słupy te są podciągane do góry dzięki ssącemu działaniu parowania (transpiracji), tworząc tzw. prąd transpiracyjny. Wraz z nim przesuwaj się w górę rośliny jony pobrane przez korzenie, a także związki organiczne, wytworzone w korzeniu, jak np. kwas glutaminowy. Szybkość prądu transpiracyjnego zależy od intensywności transpiracji; przy dużej intensywności prąd jest szybki, natomiast w przypadku ograniczenia transpiracji szybkość ruchu maleje. A zatem szybkość przewodzenia jonów i innych substancji w prądzie transpiracyjnym zależy od intensywności transpiracji, natomiast ilość substancji przetransportowanych z korzenia do części nadziemnych od transpiracji W zasadzie nie zależy zupełnie podobne jest z substancjami przewodzonymi w prądzie transpiracyjnym; jeżeli szybkość prądu zmniejszy się (np. wskutek ograniczenia transpiracji), to wzrasta wtedy stężenie substancji w pasoce i w rezultacie ilość substancji przetransportowanych w gór~ rośliny nie ulegnie większym zmianom. Dlatego intensywność transpiracji niema bezpośredniego wpływu na ilość substancji przetransportowanych do górnej części rośliny.
Uwaga końcowa Prąd transpiracyjny jest wynikiem ssącego działania parowania, Ponieważ parowanie jest procesem fizycznym, nie związanym z metabolizmem, roślina nie używa własnej energii do dalekiego transportu wody i jonów. Energia, która napędza strumień płynący wzdłuż rośliny, pochodzi więc od przyrody nieożywionej i wynika z różnicy pomiędzy siłami ssącymi gleby i atmosfery; Następstwem tej różnicy jest parowanie i transpiracja.
Daleki transport związków organicznych. Następuje on, w rurkach sitowych floemu. Transportowane są nimi przede wszystkim asymilaty od donatorów, czyli miejsca produkcji (liści) do akceptorów, czyli. miejsca zużycia (korzeni, tkanek merystematycznych, owoców)
Kierunek transportu zależy więc w pierwszym rzędzie od rozmieszczenia Iisci-donatorów. Liście umieszczone w dolnej części łodygi zaopatrują. w asymilaty głównie korzenie, natomiast liśce górnych pięter- wierzchołek wzrostu i najmłodsze liście. Ponadto .dystrybucja asymilatów zależy od innych czynników, nie zawsze znanych. Jednym z takich czynników mogą być regulatory wzrostu zwłaszcza auksyny i cytokimny
Mechanizm przewodzenia we floemie Jest to zagadnienie dyskusyjne. :Przedstawiono wiele hipotez Jedna z nich mianowicie hipoteza przepływu pod ciśnieniem jest następująca:
W procesie fotosyntezy w komórkach .mezofilu liści powstaje skrobia, ma ona jednak zbyt duże cząsteczki, by nadawała się do transportu. Ulega więc rozpadowi na mniejsze cząstki. najprawdopodobniej na glukozofosforan. Transport tego związku (bliski!)następuje w symplaście, aż do komórek towarzyszących. Tutaj heksoza, przy udziale UDPG, ulega przekształceniu na sacharozę, która z kolei zostaje aktywnie tj. przy pomocy energii metabolicznej, wydzielona do rurek sitowych. W wyniku więc aktywnego procesu. Wzrasta stężenie sacharozy w rurkach sitowych, co wywołuje osmotyczny dopływ wody i znaczny wskutek tego, wzrost ciśnienia turgorowego.
Równocześnie w innym organie rośliny (np. w korzeniu lub w bulwach) przebiegają odwrotne procesy: sacharoza jest odprowadzana z rurek sitowych (znowu ~ .udziałem energii metabolicznej !), przekształcana w komórkach towarzyszących na glukozofosforan i zużywana w procesie oddychania lub tez kierowana do amyloplastów i tu zmieniana na skrobi~. Stężenie sacharozy w rurkach sitowych wskutek tego spada powodując obniżenie ciśnienia turgorowego. A zatem ciśnienie turgorowe w układzie rurek sitowych nie jest jednakowe i przedstawia się następująco:
- w miejscu syntezy cukrów (np. w liściu) jest wysokie
- w miejscu zużycia cukrów (np. .w korzeniu) jest niskie.
Następstwem różnicy ciśnień w układzie jest masowy przepływ całej zawartości rurek sitowych, wraz ze wszystkimi rozpuszczonymi substancjami, od miejsca syntezy (wysokie ciśnienie turgorowe) do miejsca zużycia (niskie ciśnienie turgorowe).
Warunkiem funkcjonowania powyższego mechanizmu jest jednak drożność (przelotowość) sit.
Uwaga końcowa. Nie zależnie od takiej czy innej hipotezy, przewodzenie substancji we floemie jest nierozłącznie związane z oddychaniem i wydatkowaniem energii metabolicznej, uwalnianej w tym procesie. Czyli i pod tym względem można przeciwstawiać oba typy dalekiego transportu.
Przewodzenie w ksylemie (w normalnych warunkach, t!. gdy
funkcjonuje ssąca pompa transpiracji) nie wymaga dopływu energii metabolicznej, natomiast przewodzenie we floemie jest uwarunkowane jej dopływem.
CYKL HATCHA I SLACKA ( C4)
Akceptorem dwutlenku węgla w fotosyntezie jest PEP . Produktem reakcji katalizowanej przez karboksylazę PEP jest szczawiooctan potem przez redukcję powstaje jabłczan który jest regenerowany do PEP . Wynika z tego fakt że rośliny C4 mają bardzo wydajną fotosyntezę bo karboksylaza PEP odznacza się dużym powinowactwem dwutlenku węgla więc jest dużo CO2 w mezofilu przez co zmniejszona jest aktywność RuBisCO . Dodatkowo znikoma jest produkcja tlenu w chloroplastach mezofilu wieńcowego co powoduje wyeliminowanie fotorespiracji . Umożliwia to lepsze wykorzystanie nawet niskich stężeń
Floem jest podstawowym systemem przewodzącym związki organiczne na duże odległości. Transport produktów fotosyntezy składa się z 3 współzależnych od siebie etapów. Eksport asymilatów z chloroplastów do cytoplazmy ,a następnie z komórek miękiszowych liści do tkanek przewodzących to jest załadunek floemu. Następnie jest transport w samym floemie, po przemieszczeniu transportowanych związków z floemu do komórek organów - rozładunek floemu. Przewodzenie w floemie uwarunkowane jest dopływem energii metabolicznej.
SUBSTANCJE PRZEWODZONE W KSYLEMIE I FLOEMIE I MECHANIZM ICH TRANSPORTU.
Woda i sole mineralne transportowane są przez ksylem a przez floem związki organiczne w postaci wodnego roztworu ( cukry, aminokwasy , amidy, itp.) . Transport wody z solami mineralnymi:
z gleby do korzenia woda z roztworu glebowego o stężeniu osmotycznym mniejszym niż w komórkach włośników przenika do korzeni.
W korzeniu ( opory w korzeniu są dość duże bo woda z włośników przechodzi przez miękisz kory pierwotnej i endodermę aż do tkanek naczyniowych , przenikanie wewnątrz korzenia odbywa się na drodze osmotycznej)
Łodyga ( jest to najdłuższy i najtrudniejszy do pokonania odcinek dla wody . Odbywa się on dzięki siłom kohezji i adhezji , podciśnienia z transpiracji i parcia korzenia)
Transport asymilatów:
Generalnie transport ten odbywa się od miejsca produkcji asymilatów do ich zużycia lub zmagazynowania.
przenoszenie asymilatów do rurki sitowej
przepływ asymilatów
przejście asymilatów z rurki sitowej do akceptora.
OD JAKICH CZYNNIKÓW ZALEŻY WRAŻLIWOŚĆ ROŚLIN NA NIEKORZYSTNE WARUNKI ŚRODOWISKA.
Rośliny posiadają trwałe mechanizmy odpornościowe ( budowa liści, kora itp.) i nabyte - pojawiają się w odpowiedzi na czynnik stresowy. Zatem wrażliwość roślin będzie zależała od nabytych cech w toku ewolucji, czasu działania i natężenia czynnika stresowego co w ostateczności może przyczynić się do aklimatyzacji.
wrażliwości na chłód i mróz
w błonach roślin odpornych na chłód jest przewaga kwasów tłuszczowych nienasyconych.
Wyciąganie wody przez ściany komórkowe
U roślin odpornych na mróz wysokie stężenie soku komórkowego
wrażliwość i odporność na wysoką temperaturę
przewaga w błonach kwasów tłuszczowych nasyconych
ilość aparatów szparkowych umożliwiających transpirację
susza
wzrasta synteza wosków
w soku komórkowym związki osmotycznie czynne
zasolenie
odprowadzanie soli do wodniczek
związki toksyczne. Wrażliwość roślin zależy czy dany osobnik wykształcił mechanizmy przystosowawcze np. podwyższone stężenie ciężkich metali uruchamia mechanizm magazynowania nadmiaru pierwiastków w ścianach komórkowych.
10.FOTOODDYCHANIE.
W wyniku gwałtownego zaciemnienia rośliny przy fotosyntezie okazuje się że oddychanie jest bardzo intensywne. Zjawisko to wymaga wysokiego stężenie tlenu i ściśle określonej temperatury. CO2 uwalniany podczas fotooddychania ma związek RuBP. Tlen i CO2 konkurują ze sobą o miejsce w enzymie ( RuBisCO) . Przyłączenie tlenu do RuBP daje utworzenie cząsteczki PGA i jednej cząsteczki fosfoglikolanu zawierającego związany tlen. W cyklu C2 ( fotooddychanie ) 25% węgla jest tracone a procesy odbywają się przy większym udziale ATP.
FOTOODDYCHANIE - druga wersja
Fotooddychanie polega na stymulowanym przez światło wydzielaniu CO2 związanym z pobieraniem O2 . Fotooddychanie konsumuje energię metaboliczną. Fotooddychanie związane jest z właściwością karboksylazy 1,5-bisfosforybulozy , która może wiązać CO2 lub rozbijać cząsteczkę 1,5- z udziałem O2. Proces karboksylacji przewyższa ok. 2,5-3 raza utlenianie. Produktem rozszczepienia 1,5-bisfosforybulozy jest fosfoglicerynian i fosfoglikolan, który jest właściwym substratem fotooddychania .W procesie tym współdziałają chloroplasty, peroksysomy i mitochondria. Najpierw w chloroplastach następuje uwolnienie glikolanu od fosfoglikolanu. Glikolan jest transportowany do peroksysomów gdzie zostaje przekształcony w glioksalan , który następnie przekształca się w glicynę , która to przemieszcza się do mitochondriów gdzie z 2 cząsteczek glicyny powstaje 1 cząsteczka seryny oraz CO2 i NH3. Seryna następnie przemieszcza się do peroksysomów gdzie zostaje przekształcona w hydroksypirogronian a następnie w glicerynian . Glicerynian jest transportowany do chloroplastów . W fotooddychaniu następuje uwolnienie wcześniej związanego CO2 , jest więc fotooddychanie procesem przynoszącym roślinie straty Przypuszcza się, że fotooddychanie.jest procesem chroniącym aparaty fotosyntetyczne przed uszkodzeniem, gdy reakcje świetlne zachodzą z dużą wydajnością, zaś szybkość reakcji ciemnych maleje z powodu zmniejszania się zawartości CO2. Fotooddychanie zachodzi głównie w roślinach typu C3, zaś w niewielkim tylko stopniu u roślin typu C4.
11.OPIS CYKLU KALVINA BENSONA W FOTOSYNTEZIE.
W cyklu Calvina-Bensona wyróżnia się 3 fazy :
- karboksylacyjną (utworzenie 2 cząsteczek kwasu 3-fosfoglicerynowego)
- redukcyjną (polega na wytworzeniu fosfotriozy)
- regeneracyjną (polega na odtworzeniu 1,5-bisfosforybulozy)
przebieg: Karboksylaza 1,5-bisfosforybulozy przyłącza CO2 do 1,5-bisfosforybulozy dając nietrwały związek 6-węglowy, który rozpada się na dwie cząsteczki kwasu 3- fosfoglicerynowego. Kwas 3-fosfoglicerynowy jest następnie fosforylowany do kwasu 1,3-bisfosfoglicerynowego, który z udziałem NADPH ulega redukcji do poziomu cukrów , tj.do aldehydu 3-fosfoglicerynowego. Na 6 związanych cząsteczek CO2 można wyprowadzić zcyklu 2 cząsteczki trioz lub 1 cząsteczkę heksozy, które mogą być użyte do syntezy sacharozy lub skrobi, zaś pozostałe związki służą do odtworzenia akceptora dwutlenku węgla.
OPIS CYKLU KALVINA BENSONA - druga wersja
Jest to cykl zwany C3 bo produkt pierwotny fazy procesów jest trójwęglowy. Związek 3-fosfoglicerynian . Cykl można podzielić na 3 etapy:
karboksylacja rybulozo-1, 5 - bifosforanu
redukcja 3-fosfoglicerynianu
regeneracja rybulozo-1, 5 - bifosforanu
Przyłączony dwutlenek węgla do RuBP tworzy 6 węglowy nietrwały związek rozpadający się natychmiast na 2 cząsteczki PGA . Najważniejszą reakcją jest redukcja 3-fosfoglicerynianu do aldehydu 3- fosfoglicerynowego ( wykorzystuje się produkty fazy jasnej fotosyntezy NADPH i ATP) . Podstawowymi asymilatami - produktami netto fotosyntezy są skrobia i sacharoza syntetyzowane z nadwyżki fosfotrioz niewykorzystanych do regeneracji RuBP
12.PODZIAŁ BIAŁEK ENZYMATYCZNYCH.
Enzymy, czyli białka enzymatyczne są wybiórcze (w działaniu) w stosunku do różnych substancji. Dzieli się je na szereg klas:
Oxydoreduktazy - kat. reakcje ze zmianą wartościowości składników
Transferazy - kat. reakcje przenoszenia grup chemicznych między dwoma składnikami
Hydrolazy - kat. reakcje rozpadu związku z udziałem wody (esterazy, glikolazy)
Liazy - kat. reakcje odłączania grup bez udziału wody (dekarboksylazy, hydratazy )
- Izomerazy - kat. reakcje przemian wewnątrzcząsteczkowych.
Ligazy - katalizują wytworzenie wiązań między cząsteczkami w połączeniu z rozpadem związku o wiązaniach wysokoenergetycznych (potrzebują dużo energii)
(w obrębie klas wyróżniamy grupy)
13.GIBERELINA I AUKSYNA ICH ZNACZENIE PRZY RÓŻNICOWANIU WZROSTU I SŁOJÓW.
Kwas indolilo-3-octowy (IAA) jest produktem rozkładu białek. Substancja ta jest transportowana bazypetalnie z szybkością 1-2 cm/h. Auksyna jest syntetyzowana w rosnących liściach i rozwijających się wierzchołkach roślin. Auksyna powoduje wzrost wydłużeniowy komórek a w komórkach kambium wpływa „in +” na ich podzialy. Komórki wrzecionowate kambium nie mają zdolności do syntetyzowania IAA, ale tę zdolność uzyskują gdy otrzymają IAA z zewnątrz. Auksyna jest niezbędna - to jedyny hormon w obecności którego różnicują się naczynia. IAA powoduje wzrost objętościowy komórek dlatego tworzone na wiosnę cewki mają dużą średnicę. Gibereliny (np. kwas giberelinowy) także uczestniczą w regulacji wzrostu , spoczynku.
14. ZNACZENIE FE, K, Mg W METABOLIŹMIE.
Mg :
neutralizuje w komórce aniony organiczne i nieorganiczne
powoduje zmniejszenie pobierania jonów : K+, NH4+
uczestniczy w rególacji pH w komórce
wiąże się z różnymi enzymami
tworzy połączenia pomiędzy podjednostkami rybosomów
wpływa na aktywność karboksylazy rybulozobisfosforanowej w stromie chloroplastów
K :
oddziaływuje na koloidy komórkowe zwiększając ich dyspersję
neutralizuje w komórce aniony organiczne i nieorganiczne
w chloroplastach ,cytozolu, mitochondriach aktywuje ponad 50 różnych enzymów np.: ATP-azę
jego duże stężenie jest konieczne do biosyntezy białka
w błonach różnych organelli uczestniczy w kotransporcie anionów np.: NO3-
determinuje wielkość potencjału osmotycznego w komórce (osmoregulacja, ruch szparek)
występuje związek między jego stężeniem a akumulacją substancji pokarmowych
Fe :
występuje w wielu enzymach oksydoredukcyjnych i aktywuje liczne enzymy (składnik : cytochromów, ferredoksyny, peroksydazy, reduktazy azotanowej
uczestniczy w reakcjach oksydoredukcyjnych jako przenośnik elektronów
jest katalizatorem w syntezie chlorofilu i niektórych białek.
15. CO TO JEST GLIKOLIZA I JAKIE JEST JEJ ZNACZENIE W PROCESACH KATABOLICZNYCH ROŚLIN ?
Glikoliza odbywa się w cytozolu z rozkładu skrobi powstaje glukoza . Następne reakcje to utworzenie fruktozo -1, 6 - bifosforanu i jego rozłożenie, utlenienie do pirogronianu. Najważniejszym momentem glikolizy jest utlenienie aldehydu 3- fosfoglicerynowego do 3- fosfoglicerynianu. Uwalnia się energia użyta do fosforylacji i ADP , następuje redukcja NAD+. Przy przemianie 3- fosfoglicerynianu w pirogronian następuje druga fosforylacja substratowa . W sumie podczas glikolizy powstają z jednej glukozy 4 ATP ( dwie zużywane przy fosforylacji cukrów) 2NADH i dwa pirogroniany. Pirogronian przechodzi z cytozolu do mitochondrium i tam ulega dalszym przemianom- ostatecznemu utlenieniu do CO2 i H2O .
16. OMÓW REAKCJE FOTOPERIODYCZNE U ROŚLIN DRZEWIASTYCH.
Fotoperiodyzm jest to zjawisko związane z cyklicznymi dobowymi następstwami światła i ciemności ( z pieriodycznym następstwem dnia i nocy). Na fotoperiodyzm ma też wpływ zmiana długości na w stosunku do długości nocy.
Jest to zdolność do rozpoznawania i mierzenia okresów światła i ciemności, a także do rozróżniania między wydłużeniem się i skróceniem się długości dnia.
Zdolność ta jest głównym mechanizmem, który sprawia, że określone zdarzenia rozwojowe pojawiają się we właściwej porze roku.
Rośliny ze względu na reakcje fotoperiodyczne dzielimy na rośliny dnia długiego oraz dnia krótkiego oraz rośliny neutralne.
Zdarzenia rozwojowe powiązane z fotoperiodyzmem to: kwitnienie, rozmnażanie wegetatywne, tworzenie organów spichrzowych, związków zapasowych, oraz przechodzenie w stan spoczynku.
Fotoperiod jest tutaj sygnałem środowiska zewnętrznego uruchamiającym odpowiednie zmiany rozwojowe.
WERSJA DRUGA
Fotoperiodyzm jest reakcją roślin na czas trwania i periodyczne następstwo okresów światła i ciemności w cyklach dobowych. Zjawisko to opisali Garner i Allard w 1920 roku. Długość dnia i nocy zmienia się na całej kuli ziemskiej sezonowo (z wyłączeniem strefy równikowej ) dlatego też rośliny przystosowały się w znacznym stopniu do następstw występowania pór roku. Rośliny kwitną zależnie od faz jasnej i ciemnej w 24 godzinnym cyklu dobowym. Wyróżniamy następujące grupy fotoperiodyczne:
rośliny dnia krótkiego SDP kwitnące gdy w dziennym fotoperiodzie jest przewaga fazy ciemnej ( powyżej 12 godzin ciemności).
Rośliny dnia długiego LDP kwitnące gdy w dziennym fotoperiodyźmnie jest przewaga fazy jasnej ( powyżej 12 godzin jasnośći)
Rośliny dnia krótkiego- długiego. SLDP i długiego - krótkiego LSDP . Ich inicjacja zależy od ekspozycji na dzień krótki i długi w odpowiedniej kolejności
Rośliny neutralne - DNP . Niewrażliwe na długość dnia. W przypadku tej grupy roślin sygnałem inicjacji kwitnienia jest osiągnięcie przez roślinę odpowiedniego wieku bądź wielkości.
Reakcje fotoperiodyczne:
ilościowa reakcja fotoperiodyczna tzw. wrażliwość względna. Ekspozycja na indywidualny fotoperiod tylko przyspiesza lub hamuje kwitnienie.
Jakościowa reakcja fotoperiodyczna -wrażliwość bezwględna- ekspozycja na odpowiedni fotoperiod bezwzględnie warunkuje indukcję fotoperiodyczną . Niektóre rośliny wymagają tylko jednego indukcyjnego fotoperiodu. Inne wymagają kilku stosownych fotoperiodów.
17. MECHANIZM TRANSPORTU BIERNEGO I CZYNNEGO
W zależności od rodzaju transportowanej substancji oraz czynników warunkujących pojawienie się transportujących te substancje przez błony wyróżniono kilka sposobów transportu :
Transport bierny - polega na przemieszczaniu się substancji w obu kierunkach ale zawsze zgodnie z gradientem stężeń. W ten sposób gazy (tlen, azot, CO2, NH3) i mocznik oraz niektóre jony dyfundują wraz z prądem wody przez błonę do miejsc mniejszym stężeniu. Transport bierny zachodzi do czasu wyrównania się różnicy stężeń po obu stronach błony.
Dla cząsteczek związków organicznych i niektórych jonów (K+, Cl- ) błony stanowią przeszkodę, którą mogą pokonać tylko przy udziale nośników (białek nośnikowych -ruchliwych składników błon). Jeżeli substancja jest transportowana przez nośnik zgodnie z gradientem stężeń to proces taki nie wymaga nakładu energii i nazywamy go transportem ułatwionym lub ułatwioną dyfuzją.
Transport aktywny wymaga obecności nośników i dopływu energii pochodzącej z przemian metabolicznych. W ten sposób następuje przemieszczanie się nie tylko wielu jonów, lecz również związków organicznych, np. cukrów lub aminokwasów, wbrew gradientowi stężeń. Transport przy udziale nośników jest zwykle jednokierunkowy. Ten typ transportu substancji jest niezbędny do życia komórki ponieważ warunkują pobieranie przez nią składników koniecznych do prawidłowego funkcjonowania nawet wtedy, gdy ich stężenie w środowisku jest bardzo niskie, wówczas wymaga nakładu energii.
Transport cieczy i cząsteczek większych od tych, które mogą być przemieszczane przez pory błon lub przy udziale nośników odbywa się na drodze endocytozy - przenikania do wnętrza komórki wyniku tworzenia się wakuol. Pobieranie cieczy i koloidów następuje w procesie pinocytzy a ciał stałych przez fagocytozę. Fagocytoza wymaga dostarczenia energii. Wypieranie różnych substancji z komórki do środowiska zachodzi w procesie egzocytozy. W ten sposób wydzielane są np. hormony i enzymy oraz produkty przemiany materii.
18.CO OZNACZA TERMIN STRES W FIZJOLOGII ROŚLIN.
Stres to stan pewnego „napięcia” w organizmie w którym wskutek działania niesprzyjających czynników środowiska następuje upośledzenie albo ograniczenie metabolizmu lub rozwoju . Czynnikiem stresowym może być : zbyt niska lub wysoka temperatura , brak lub nadmiar wody , nadmierne zasolenie lub brak soli , światła itp. Rośliny wykształciły mechanizmy obronne przed działaniem czynników stresowych. Są to mechanizmy trwałe i wykształcone ( aklimatyzacja)
19.CYKL KREBSA - PRZEBIEG I ZNACZENIE.
Reakcje cyklu odbywają się w mitochondrium. AcetyloCoA zostaje przyłączony do szczawiooctanu i powstaje 6 węglowy cytrynian potem izomer izocytrynian. Przy późniejszej reakcji odłączenia CoA uwalnia się energia umożliwiająca syntezę ATP na drodze fosforylacji . Dalsza reakcja to utlenienie bursztynianu . Elektrony nie są przenoszone na NAD+ ale na FAD powstaje FADH2 . Fumaran ( utleniony bursztynian jest przekształcony w jabłczan który jest utleniany na szczawiooctan przy czym NAD NADH . W ten sposób cykl zostaje zamknięty . Z jednej cząsteczki pirogronianu powstają 3 cząsteczki CO2 i cztery NADH i jedna NADH2 i jedna ATP. Niecały węgiel zostaje utleniony do CO2 dalej może on służyć w reakcjach syntezy innych substancji ( alanina, kwasy tłuszczowe , cytochromy)
CYKL KREBSA - PRZEBIEG I ZNACZENIE. - druga wersja
Cykl Krebsa znany też jako cykl kwasu cytrynowego to proces w którym pochodzący od pirogronianu octan zostaje utleniony do CO2 i H2O. Powstają przy tym kolejne cząsteczki NADH i jedna FADH 2 . Enzymy tego cyklu znajdują się w macierzy mitochondrialnej ( matrix) i tam odbywają się reakcje cyklu. Głównym substratem cyklu Krebsa jest pirogronian , który najpierw jest przetransportowany z cytoplazmy do wnętrza mitochondrium - w matrix pirogronian ulega karboksylacji do acetylokoenzymu A. Acetylokoenzym A w poszczególnych ogniwach cyklu Krebsa ulega :
dwukrotnej dekarboksylacji z wydzieleniem dwóch cząsteczek CO2 oraz czterokrotnej dehydrogenacji z wydzieleniem 3 cząsteczek NADH i jednej FADH2
Poszczególne etapy cyklu Krebsa:
przyłączenie grupy acetylo-CoA do szczawiooctanu i powstanie cytrynianu.
Cytrynian przechodzi w izocytrynian który w procesie dekarboksylacji daje oksoglutaron, NADH i CO2
Oksoglutaron podlega także procesowi dekarboksylacji, przyłącza się znów acetylo- CoA i powstaje sukcynylo- CoA, NADH i CO2
Sukcynylo-CoA przekształca się w bursztynian, uwalnia się wtedy energia umożliwiająca syntezę ATP na drodze fosforylacji substratowej
Bursztynian jest utleniany na fumaron. Powstaje wtedy FADH2
Fumaron przekształca się w jabłczan.
Jabłczan utleniany jest w szczawiooctan i jednocześnie powstaje cząsteczka NADH.
Zregenerowany szczawiooctan zdolny jest do reakcji z kolejnym acetylo-CoA. W ten sposób cykl przemian zostaje zamknięty i może rozpocząć się od nowa.
Znaczenie cyklu:
- w cyklu tym następuje utlenienie cukrów a także tłuszczów i białek przekształconych na czynny octan.
- jest też ważnym dostawcą metabolitów do różnych syntez
-stanowi pulę różnych łańcuchów węglowych substratów do różnorodnych procesów biochemicznych w komórkach roślinnych
- w tkankach niefotosyntetyzujących jest jedynym źródłem ATP i zredukowanych nukleotydów do syntezy nowych związków.
20.INHIBITORY WZROSTU.
ETYLEN - jest to związek o bardzo prostej budowie. Substratem w syntezie jest metionina. Etylen hamuje wzrost wydłużeniowy siewek, przyspiesza procesy dojrzewania, starzenia się tkanek i warstwy odcinającej u liści i owoców.
KWAS ABSCYSYNOWY (ABA) - związek o strukturze przypominającej karotenoidy. Jego synteza odbywa się z kwasu mewalonowego lub pośrednio z karotenoidów. Wytwarzany jest w komórkach zawierających plastydy w organach pod działaniem stresu. Indukuje stan spoczynku w pąkach i w nasionach gdzie jest inhibitorem kiełkowania. Hamuje wzrost objętościowy i wydłużeniowy komórek. Hamuje fotosyntezę , syntezę chlorofilu i transport jonów przez błony, indukuje zamykanie się aparatów szparkowych, przyspiesza starzenie
JASMONIANY- kwas jasmonowy przypomina strukturą ABA . Hamują wzrost elongacyjny pędu , kiełkowanie pyłku i nasion oraz tworzenie się pąków pędowych i kwiatowych. Stymuluje zamykanie się aparatów szparkowych , rozwój bulw , dojrzewanie owoców, rozkład chlorofilu, starzenie się liści. Podobnie jak ABA towarzyszą reakcji roślin na stresy.
ZWIĄZKI FENOLOWE- stanowią wielką grupę substancji produkowanych przez rośliny większość syntetyzowana jest z aminokwasu fenyloalaniny. Są to inhibitory wzrostu i kiełkowania mają udział w allelopatii. Ogólnie nie mają cech typowych hormonów
21.CO TO JEST FIZJOLOGIA ?
Fizjologia roślin jest to dyscyplina botaniki zajmująca się badaniem procesów zachodzących w organizmach roślin, a zwłaszcza biochemicznych i biofizycznych oraz procesów rozwoju i wzrostu od zapłodnienia do śmierci. To wiąże fizjologię z embriologią, ekologią roślin, biochemią i biofizyką.
Fizjologia bada procesy zachodzące na poziomie ponadkomórkowym: poziom tkankowy, poziom organizmu; oraz na poziomie subkomórkowym: poziom organelli, poziom ponadmolekulalny, poziom molekulalny, poziom submolekulalny.
Fizjologia wyjaśnia procesy, w których rośliny przekształcają proste surowce - dwutlenek węgla, wodę i sole mineralne w skomplikowane substancje zaspokajające potrzeby ludzi. Znajomość tych procesów pozwala ulepszać metody uprawy roli i roślin. Badania nad fizjologią żywienia mineralnego są wykorzystywane przy doskonaleniu metod nawożenia. Osiągnięcia fizjologii wzrostu i rozwoju przyczyniają się do wprowadzania nowoczesnych metod regulacji plonów. Praktycznym zastosowaniem osiągnięć fizjologii roślin są chemiczne regulatory wzrostu. Stosowanie ich pozwala zwiększyć siłę i energię kiełkowania, sztucznie ukorzeniać sadzonki, regulować terminy kwitnienia, wpływać na zawiązywanie, dojrzewanie i opadanie owoców, selektywnie tępić choroby w zasiewach itp.
22. ZNACZENIE TRANSPIRACJI.
Transpiracja wywiera na rośliny zarówno dodatni jak i ujemny. Dodatnie oddziaływanie utraty wody przez wyparowanie sprowadza się do tego, że:
Transpiracja jest niezbędna do uruchomienia głównego (kohezyjnego) mechanizmu pobierania i transportu wody w roślinie, jakkolwiek przy braku tego procesu dostawa wody do wszystkich części rośliny jest zapewniona za pośrednictwem parcia korzeniowego.
Ułatwia chociaż w ograniczonym rozmiarze, pobieranie składników mineralnych z gleby. Nie należy przeceniać roli transpiracji w tym procesie, gdyż istnieje duża niezależność pomiędzy tempem utraty wody, a pobieraniem jonów z roztworu glebowego.
Obniża temperaturę organów transpiracyjnych w wyniku zużywania energii cieplnej w formie ciepła parowania, co często chroni roślinę przed przegrzaniem. Obniżenie temperatury liści w wyniku transpiracji jest na ogół niewielkie i wynosi 1-4 C. Proces ten nie jest jedynym sposobem utraty ciepła przez roślinę. Większe ubytki energii cieplnej zachodzą przez wypromieniowanie oraz przez konwencję, gdy temperatura liścia jest wyższa niż temperatura powietrza.
Intensywna transpiracja wywołująca okresowe wahania w zawartości wody w roślinie, a tym samym dobowe zmiany w turgorze komórek wpływa na wzrost liści, pędów i owoców. Znaczny spadek turgoru komórek wywołuje z reguły ograniczenie tempa wzrostu tych organów , co jest korzystne ze względu na lepsze wykorzystanie tkanek wzmacniających. Lepsze wykształcenie tkanek mechanicznych, gruba kutykula i grubsze ściany komórkowe zwiększają ponadto odporność roślin na czynniki chorobotwórcze.
Poza wymienionymi dodatnimi stronami transpiracji, proces ten uważa się za zło konieczne, powodujące więcej uszkodzeń i przypadków nieodwracalnego (letalnego) więdnięcia roślin, niż jakikolwiek inny czynnik niekorzystnie działający na rośliny.
23. ZNACZENIE PROCESÓW REDOKS W PROCESIE METABOLIZMU.
Reakcja utleniania polega na odbieraniu elektronów od utlenionej cząsteczki przez tlen lub inne cząsteczki łatwo przyjmujące elektrony. Koleje ogniwa łańcucha oddechowego charakteryzują się zdolnością do coraz łatwiejszego przyjmowania elektronów. Są więc one uszeregowane według wzrastającego powinowactwa do elektronów, czyli według wzrastających potencjałów oksyredukcyjnych. Dzięki temu wzdłuż tego łańcucha płynie strumień elektronów lub atomów wodoru.
Komórki uzyskują energię jedynie w wyniku ruchu elektronów. Ma to właśnie miejsce w procesach oksyredukcyjnych, np. podczas przepływu elektronów przez łańcuch oddechowy albo przez łańcuch transportu elektronów w procesie fotosyntezy. Dlatego wszystkie przemiany oksyredukcyjne ( np. oddychanie i fotosynteza )są nierozłącznie związane z przekształceniami energii.
Zestaw koenzymów i enzymów zwany łańcuchem oddechowym tworzy jak gdyby szczeble drabiny, po których opuszczają się elektrony, podczas biologicznego utleniania. Na poszczególnych szczeblach wydzielają się określone porcje energii przechwytywanej i zużywanej na reakcję:
A D P + Pi + energia ATP
(adenozyno- (fosforan (adenozynotrójfosforan) --> [Author:TMH] dwufosforan) nieorganiczny)
Energia strumienia elektronów przepływającego przez łańcuch oddechowy zostaje zatem zmagazynowana w makroenrgetycznych wiązaniach fosforowych ATP. Proces ten nazywamy fosforylacją w łańcuchu oddechowym.
Łańcuch transportu elektronów w procesie fotosyntezy jest wbudowany w membrany tlakoidów. Na początku każdego łańcucha transportu stoi donor czyli związek przejawiający tendencję do oddawania elektronów (aktywy chlorofil P700 lub P680). Elektrony oddane przez donor zostają przekazane na akceptor. Akceptor przekazuje elektrony na następny akceptor, ten znów na następny itd.
Podczas przepływu przez kolejne ogniwa łańcucha transportu elektrony stopniowo tracą swą energię. Energia ta nie ginie lecz jest przekazywana i wiązana chemicznie w postaci fosforanowych wiązań ATP. Przepływ strumienia elektronów przez łańcuch przenośników, wywołany energią świetlną jest sprzężony z syntezą wysokoenergetycznych fosforanów typu ATP. Proces ten nazywamy fosforyzacją fotosyntetyczną.
ADP +Pi ATP
Donor elektronu akceptor elektronu
24. CO TO JEST POTENCJAŁ WODY I JAKIE JEST JEGO ZNACZENIE ?
O przechodzeniu wody z jednej komórki do drugiej decyduje różnica pomiędzy ich siłami ssącymi, co wyraża się równaniem S=W-T, gdzie W-oznacza wartość osmotyczną soku komórkowego, a T ciśnienie turgorowe komórki.
Jednakże w ujęciu ogólnym siła ssąca komórek nie polega wyłącznie na zjawiskach osmotycznych, gdyż w grę wchodzą także: pęcznienie różnych substancji, a zwłaszcza biokoloidów, adchezja itp. W celu całościowego rozumienia stanu uwodnienia komórek, tkanek i organizmów roślinnych należy oprzeć się na termodynamicznym pojęciu potencjału wody.
O wielkości potencjału wody stanowi stosunek różnicy między wartością potencjału chemicznego wody danego roztworu lub danej substancji i potencjału chemicznego czystej wody do cząstkowej objętości molalnej wody w rozpatrywanym układzie:
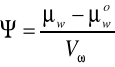
μw-potencjał chemiczny wody roztworu
μwo- potencjał chemiczny czystej wody
Vw - cząstkowa objętość molalna wody układu
Chemiczny potencjał wody można rozumieć intuicyjnie jako stan nią w danym układzie. Jest on miarą zdolności danej substancji do wykonywana pracy. Wartość potencjału chemicznego czystej wody pod ciśnieniem 1 atmosfery przyjmuje się za 0; stąd Ψ ma wartość ujemną.
Przepływ wody pomiędzy dwoma układami skierowani jest do bardziej negatywnej wartości Ψ(czyli do układu mniej uwodnionego w stosunku do jego możliwości ).
W układach nieożywionych Ψwykazuje przy normalnym ciśnieniu wartość ujemną, to jednak w organizmach żywych nie zawsze tak musi być. Przykładem mogą być gutujące rośliny, które „aktywnie” wydzielają wodę.
Potencjał wody wyrażamy w jednostkach ciśnienia.
25. OD CZEGO ZALEŻY SZYBKOŚĆ REAKCJI ENZYMATYCZNEJ ?
Na szybkość reakcji katalizowanej przez enzym wpływa wiele czynników, a mianowicie:
a) temperatura
b) pH środowiska reakcji
c) stężenie enzymu
d) stężenie substratu
Wraz ze wzrostem temperatury do 30-37°C szybkość reakcji enzymatycznej wzrasta, osiągając maksimum w temperaturze optymalnej. W wyższej temperaturze enzym stopniowo traci sprawność działania w wyniku postępujących zmian w strukturze przestrzennej białka, prowadzących do jego unieczynnienia. Amfoteryczny charakter aminokwasów i białek sprawia, że enzymy działają w pewnych zakresach pH ujawniając maksymalną sprawność katalitycznego działania przy ściśle określonym stężeniu jonów wodorowych w środowisku reakcji. Optimum pH dla większości enzymów roślinnych mieści się w granicach 4,5-7,5. Wiele enzymów katalizuje przebieg reakcji w obydwu kierunkach. W takich przypadkach każdy z kierunków reakcji charakteryzuje się odmiennym optimum pH.
pH 7,5
jabłczan fumaraza fumaran +H2O
pH 6,2
Wzrost stężenia substratu przyśpiesza reakcję, która osiąga maksymalną wydajność wtedy, gdy wszystkie cząsteczkienzymu są połączone z substratem. Gdy w komórce znajdują się tylko kompleksy E-S (enzym-substrat)to szybkość reakcji jest stała i osiąga maksymalną wartość, a dalszy wzrost stężenia substratów doprowadza do wyhamowania procesu
Wykresy obrazujące zależność szybkości reakcji od następujących czynników :a)temperatury; b)pH; c)stężenia enzymu; d)stężenia substratu
a) b)
Temp. opt Temp. optimum PH
d) Szybkość reakcji
Szybkość reakcji
stężenie stężenie
enzymu substratu
26. MIEJSCE POWSTAWANIA I PROCES PRZEMIESZCZANIA SIĘ STYMULATORÓW WZROSTU.
Do regulatorów wzrostu zaliczamy:
auksyny - powstają w zielonych częściach roślin w formie nie czynnej jako tzw. proauksyny i stąd rozprowadzane są do części wzrostowych, gdzie przetworzone zostają na formy aktywne. Ze stożków wzrostu spływają w dół floemem. W części nadziemnej rośliny auksyna jest transportowana od wierzchołka pędu do podstawy - jest to transport bazypetalny; natomiast w korzeniu auksyna jest transportowana od podstawy rośliny do wierzchołka korzenia-akropetalny.
gibereliny- wytwarzane są głównie w liściach, skąd spływają w dół floemem. Z korzeni mogą „wędrować” w górę ksylemem. Transport giberelin nie ma więc charakteru polarnego.
cytokininy- wytwarzane są głównie w stożkach wzrostu u korzeni, skąd wędrują w górę z prądem transpiracyjnym (ksylemem).
inhibitory wzrostu - np. kwas obscysynowy, kwas jasmonowy. Kwas obscysynowy (ABA) powstaje w różnych organach, szczególnie zaś w starych liściach - miejscem syntezy są plastydy. Transportowany jest głównie floemem, ale także miękiszem oraz ksylemem.
27.TRANSPORT ELEKTRONÓW I ZNACZENIE W METABOLIŹMIE
Elektrony wędrują z H2O na NADP+ przez łańcuch przenośników. Jest to szereg układów oksydoredukcyjnych , których większość została wyizolowana i scharakteryzowana. Transport elektronów w fazie świetlnej fotosyntezy odbywa się z udziałem 3 dużych kompleksów białkowych. Są to fotosystem II, kompleks cytochromowy b6-f i fotosystem I . PSII po utracie elektronów nabiera silnych właściwości utleniających pozwalających na utlenienie wody , a pochodne chlrofilu a 680 (P680) elektrony przenoszone są za pośrednictwem plastochinionu na kompleks cytochromowy b6-f . Z kompleksu cytochromów przechodzą na PS I za pośrednictwem plastocyjaniny. Wybicie elektronu z chlorofilu a700 ( P700) powoduje pojawienie się właściwości utleniających pozwalających na utlenienie cytochromów oraz silnych zdolności redukcyjnych prowadzących do redukcji NADP+. Podczas wędrówki elektronu następują stopniowe zmiany potencjałów oksydoredukcyjnych kolejnych przenośników. Występuje tu paradoksalna sytuacja, gdzie układ o wysokim potencjale dodatnim redukuje układ o wysokim potencjale ujemnych. Redukcja taka wymaga dużego nakładu energii. Fotosysem II (PS II) działający z maksymalną efektywnością przy 680 nm długości fali światła zlokalizowany jest głównie w tylakoidach gran. Wybicie elektronów z centrum reakcji PS II powoduje odciągnięcie elektronu z H2O co wiąże się z wydzieleniem O2 i uwolnieniem protonów ( H+). Przyjęcie elektronu z H2O powoduje powrót P680 do formy zredukowanej, a elektron wybity przedtem przez foton zostaje przekazany na feofitynę ( związek podobny do chlorofilu). Z feofityny elektrony przechodzą na plastochiniony. Ostatni z nich przyjmując elektron pobiera ze stromy protony i przekształca w hydroplastochiniony. W tej postaci zanurza się w błonie i przekazuje elektrony na układ cytochromowy, a protony uwalnia do wnętrza tylakoidu.
Mechanizm wędrówki elektronu przez kompleks cytochromowy nie jest dokładnie poznany. Fotosytem I czyli PSI występuje głównie w tylakoidach stromy. Centrum reakcji PS I po wybiciu z niego elektronu przez kolejny foton staje się słabym utleniaczem odbierającym elektron od plastocyjaniny, która łącznie z plastochinionem pełni funkcję ruchliwego łącznika między PS II a PS I . Równocześnie PS I staje się silnym reduktorem oddającym elektrony na kolejne przenośniki. Pierwsze układy pośredniczące w przekazywaniu elektronu z PS I są bardzo nietrwałe. Znane są natomiast następne, są to żelazo-siarkowe białka które przenoszą elektrony na ferrodoksynę. W końcu flawoproteinowy enzym przenosi elektron na NADP+ i redukuje go do NADPH. NADPH stanowi końcowy produkt transportu elektronu z H2O. Przepływ elektronu od PS II przez PS I do NADP+ jest to tzw. niecykliczny transport elektronu. Może on być uzupełniany przez przepływ cykliczny w którym ferrodoksyna przekazuje elektron z powrotem na układ cytochromowy b6 -f, a następnie przy współudziale plastochinionu na PS I . Przy okazji zostają do kanału tylakoidowego przeniesione dodatkowe protony. Znaczenie tej wędrówki elektronu jest ogromne. Powstaje bowiem nie tylko NADPH ale również ATP. Są to podstawowe związki wysokoenergetyczne wykorzystywane do xxxxxxxxx i redukcji CO2 i wytwarzania substancji organicznych. Bez tego przepływu nie istniała by fotosynteza.
Transport elektronów i znaczenie w metaboliźmie- wersja druga
Następujące przenośniki, odpowiednio uszeregowane, tworzą łańcuch transportu elektronów, który jest wbudowany w membrany tylakoidów:
Ferredoksyna (białko zawierające siarkę oraz żelazo)
Plastocyjanina (białko w którym występuje miedź )
Plastochinon (związek zbudowany z aromatycznego pierścienia i bocznego łańcucha )
Cytochromy b i f (zasadniczą częścią cząsteczki jest pierścień porfirynowy zawierający w centrum atom żelaza)
Nukleotydy nikotynoamidoadeninowe (związki typu NADP i NAD)
Na początku łańcucha transportu elektronów stoi donor ( związek przejawiający tendencję do oddawania elektronów). W procesie fotosyntezy donorem elektronów jest aktywny chlorofil P-700 lub P-680. Elektrony oddawane są na akceptor, czyli związek przejmujący elektron. Każdy przenośnik jest donorem elektronów. Przepływ elektronów wiąże się z rosnącym powinowactwem przenośników ( potencjałem oksydoredukcyjnym). Elektrony dzięki swemu ruchowi zawsze maja energię. Najwięcej energii mają elektrony uwalniane w skutek pochłoniętej z chlorofilu energii świetlnej. Elektrony tracą swą energię podczas przepływu przez kolejne ogniwa łańcucha przenośników. Energia ta jest przechwytywana i wiązana chemicznie w postaci ATP. Transport elektronów wiąże się więc z synteza ATP. Proces ten nazywany jest fosforylacją fotosyntetyczną.w zależności od układu przenośników elektronów wyróżniamy dwa typy fosforylacji fotosyntetycznej:
fosforylacja fotosyntetyczna niecykliczna. System barwników PSI (chlorofil P-700) pochłania fotony światła ciemnoczerwonego. Energia pochłoniętego fotonu, przekazana do aktywnego centrum, powoduje wybicie elektronu o dużym zapasie energii. W miejscu wybitego elektronu powstaje „dziura”, która może przyjąć elektron z zewnątrz. Uwolniony elektron jest przekazywany przez kolejne przenośniki:A0, A1, Fx ,FB , FA na ferredoksynę, a z niej na dwunukleotyd NADP. Wodór, pochodzący z fotolizy wody, zostaje również skierowany na cząsteczkę NADP która ulega przez to redukcji i powsaje NADPH2 , czyli pierwszy zasadniczy składnik „siły asymilacyjnej”. Tymczasem jony OH-, pochodzące z cząsteczki wody ulegają reakcji, co powoduje uwalnianie się tlenu, wydalanego przez roślinę jako jeden z końcowych produktów fotosyntezy. Oba elektrony (2e-) zostają skierowane na system barwników PSII (chlorofil P-680), gdzie wypełniają „dziury”, a następnie są wybijane pod wpływem światła jasnoczerwonego. Wyzwolone elektrony, również o dużym zapasie energii przepływają przez łańcuch przenośników z powrotem ma system PSI i w ten sposób wypełniają „dziury” powstałe poprzednio w tych barwnikach. Po pochłonięciu nowych fotonów zespół PSI może ponownie wysyłać w obieg wybite elektrony
Fosforylację fotosyntetyczną niecykliczną można przedstawić w postaci sumarycznego równania:
światło
NADP + H2O + ADP + Pi NADPH2 +ATP+1/2 O2
Chloroplast
fosforylacja fotosyntetyczna cykliczna elektrony wybite pod wpływem światła z systemu barwników PSI przepływają przez łańcuch przenośników z powrotem na system PSI z pominięciem NADP. Energia, uwolniona podczas tego przepływu, zostaje zmagazynowana w postaci ATP. W procesie fosforylacji cyklicznej nie następuje zatem fotoliza wody i powstaje tylko część „siły asymilacyjnej”, gdyż nie tworzy się NADPH2.. sumaryczne równanie tego procesu:
Światło
ADP + Pi ATP
Chloroplast
Znaczenie w metabolizmie
Fosforylacja fotosyntetyczna cykliczna i niecykliczna są częściami fazy jasnej procesu fotosyntezy, która jest typowym procesem anabolicznym, czyli powstałe w niej związki znajdują się na wyższym poziomie energetycznym niż substraty biorące udział w reakcji. W obu typach fosforylacji powstaje ATP, który stanowi akumulator energii i jest zasadniczym składnikiem „siły asymilacyjnej”
WPŁYW FITOCHROMÓW NA ODPORNOŚĆ ROŚLIN , NA NIEKORZYSTNY WPŁYW ŚRODOWISKA.
Fitochrom jest to fotoreceptor uwalniany w reakcjach fotoperiodycznych ( reakcja roślin na względną długość kolejno zmieniających się okresów świetlnych i ciemności) i wielu innych wywołanych przez światło fizjologicznych reakcjach roślin. Jest to niebiesko zielony barwnik o charakterze białkowym . Występuje u wszystkich roślin naczyniowych w dwóch postaciach - formach łatwo ulegających wzajemnym przekształceniom w wyniku absorbcji światła ( fotokonwersji).
I PR lub P660 - silnie pochłania czerwone światło o długości fali 660 nm.
II PFR lub P 730 - silnie pochłania czerwone światło o długości fali 730 nm.
( tzw. daleka czerwień)
PFR jest to postać mało stabilna i przekształca się samorzutnie PR. Jest to jednak czynna forma wywołująca reakcje kwitnienia. W roślinie jest mieszanina PFR i PR ale przeważa PFR ( dzień) a w nocy PFR powoli przekształca się w PR . To pozwala orientować się roślinie w środowisku i dobrać porę kwitnienia i wydawania potomstwa w warunkach korzystnych dla danego gatunku roślin. Rośliny które wymagają do rozwoju i rozmnażania wysokich temperatur będą kwitnąć gdy okres nasłonecznienia będzie wysoki co uchroni je przed wyginięciem ich potomstwa np. od mrozu czy niekorzystnych innych warunków atmosferycznych.
Dzień krótki - PFR hamuje kwitnienie, długa noc powoduje że rośliny w pewnym okresie nie zawierają w ogóle PFR i następuje inicjacja kwitnienia. Wystarczy długą noc przerwać 10 minutami wystawienia na światło aby rośliny takie nie zakwitły. PR przekształca się w PFR. Jeżeli jednak taką roślinę ponownie podda się działaniu krótkiego błysku dalekiej czerwieni to roślina zakwitnie
Gdy PFR absorbuje daleką czerwień przekształca się na powrót w formę wyjściową PR.
Dzień długi- długie dni - produkcja głównie PFR krótka noc- pewne ilości PFR przekształcają się w PF ale pozostaje dość PFR aby mogła zajść indukcja kwitnienia. PFR indukuje kwitnienie roślin dnia długiego.
Fitochrom bierze także udział w kontroli kiełkowania nasion, ruchach sennych liści, spoczynku pędów, opadaniu liści oraz powstawaniu barwników w kwiatach, owocach i liściach.
Wpływ fitochromów na odporność roślin , na niekorzystny wpływ środowiska.- wersja druga
Fitochrom jest to barwnik, który uczestniczy w rejestrowaniu zmian zachodzących w środowisku wzrostu roślin i dostarcza im informacje o tych zmianach. Zmiany te dotyczą głównie modyfikacji widma słonecznego w zakresie światła czerwonego i dalekiej czerwieni. Obecność fitochromu stwierdzono w komórkach i tkankach wszystkich roślin, ale nie grzybów. Barwnik ten funkcjonuje w dwóch formach molekularnych różniących się między sobą właściwościami spektralnymi: Pr wykazuje maksimum absorpcji w zakresie światła czerwonego, a Pfr pochłania światło dalekiej czerwieni. Powszechnie przyjmuje się, że jedynie Pfr jest aktywną formą fitochromu. Naświetlanie roślin światłem czerwonym lub dalekiej czerwieni towarzyszy ustalenie się tzw. stanu fotostacjonarnego, który decyduje o powstaniu określonej odpowiedzi fotomorfogenetycznej. Pochłonięciu światła towarzyszy uruchomienie skomplikowanego łańcucha przemian metabolicznych, zwanego również łańcuchem przekazywania sygnału, którego przebieg zależnie od stanu kompetencji komórki prowadzi do ujawnienia się adekwatnej odpowiedzi komórkowej.
Istotną rolę w powstawaniu odpowiedzi morfogenetycznej odgrywa dawka światła pochłoniętego przez fitochrom. W związku z tym procesy regulowane przez ten fotoreceptor można podzielić na trzy kategorie:
Reakcje bardzo niskoenergetyczne (do zaindukowania reakcji wystarczy powstanie znikomej około 1% ilości fitochromu aktywnego)
Reakcje niskoenergetyczne (wymaga większej ilości aktywnego fitochromu )
Reakcje wysokoenergetyczne (wymagają długiego lub cyklicznie powtarzanego okres światła)
Fitochrom rejestrując zmiany zachodzące w środowisku wzrostu rośliny dostarcza jej informacji o niekorzystnych dla niej czynnikach. Dzięki takiej reakcji wpływa na powstawanie odporności rośliny na niekorzystne warunki środowiska.
Wpływ fitochromów na odporność roślin , na niekorzystny wpływ środowiska.- wersja druga
Fitochrom jest to barwnik, który uczestniczy w rejestrowaniu zmian zachodzących w środowisku wzrostu roślin i dostarcza im informacje o tych zmianach. Zmiany te dotyczą głównie modyfikacji widma słonecznego w zakresie światła czerwonego i dalekiej czerwieni. Obecność fitochromu stwierdzono w komórkach i tkankach wszystkich roślin, ale nie grzybów. Barwnik ten funkcjonuje w dwóch formach molekularnych różniących się między sobą właściwościami spektralnymi: Pr wykazuje maksimum absorpcji w zakresie światła czerwonego, a Pfr pochłania światło dalekiej czerwieni. Powszechnie przyjmuje się, że jedynie Pfr jest aktywną formą fitochromu. Naświetlanie roślin światłem czerwonym lub dalekiej czerwieni towarzyszy ustalenie się tzw. stanu fotostacjonarnego, który decyduje o powstaniu określonej odpowiedzi fotomorfogenetycznej. Pochłnięciu światła towarzyszy uruchomienie skomplikowanego łańcucha przemian metabolicznych, zwanego również łańcuchem przekazywania sygnału, którego przebieg zależnie od stanu kompetencji komórki prowadzi do ujawnienia się adekwatnej odpowiedzi komórkowej.
Istotną rolę w powstawaniu odpowiedzi morfogenetycznej odgrywa dawka światła pochłoniętego przez fitochrom. W związku z tym procesy regulowane przez ten fotoreceptor można podzielić na trzy kategorie:
Reakcje bardzo niskoenergetyczne (do zaindukowania reakcji wystarczy powstanie znikomej około 1% ilości fitochromu aktywnego)
Reakcje niskoenergetyczne (wymaga większej ilości aktywnego fitochromu )
Reakcje wysokoenergetyczne (wymagają długiego lub cyklicznie powtarzanego okres światła)
Fitochrom rejestrując zmiany zachodzące w środowisku wzrostu rośliny dostarcza jej informacji o niekorzystnych dla niej czynnikach. Dzięki takiej reakcji wpływa na powstawanie odporności rośliny na niekorzystne warunki środowiska.
29.ROLA CZYNNIKÓW HORMONALNYCH W PROCESACH REGENERACJI ROŚLIN DRZEWIASTYCH.
Hormony roślinne to inaczej regulatory wzrostu, do których zaliczamy: auksyny, gibereliny, cytokininy, inhibitory wzrostu, etylen.
Hormonem roślinnym wpływającym na regenerację roślin drzewiastych jest auksyna. Auksyna wydłuża komórki, wywołuje ich podziały. Jednym z przykładów pobudzania podziałów komórkowych przez ten hormon jest powstawanie kallusa. W momencie zranienia rośliny komórki zranionego miejsca dzielą się intensywnie wytwarzając luźną parenchymatyczną tkankę- kallus, która zarasta uszkodzone miejsce. Posmarowanie zranionego miejsca pastą lub roztworem zawierającym auksynę pobudza podziały komórkowe co wpływa na zwiększenie kallusa. Zjawisko to może być wykorzystywane w leczeniu ran u roślin, oraz w przyspieszaniu zrostu zrazu z podkładką przy zabiegu szczepienia.
30. ZNACZENIE GIBERYLINY W PROCESIE WZROSTU I RÓŻNICOWANIA DRZEW.
Giberelinę - hormon roślinny odkrył w latach 20-tych naszego wieku japoński badacz Kurosawa. Z grzyba wywołującego chorobę „wściekłego ryżu” wyizolował on krystaliczną, aktywną substancją zwaną gibereliną. Największe stężenie giberelin występuje w młodych, rozwijających się nasionach i owocach, które stanowią miejsce ich syntezy. Gibereliny są tworzone w wierzchołkowych partiach pędu oraz w korzeniu. Główna, aktywna biologicznie giberelina u roślin to giberelina A1.. Podstawowym przejawem działania giberelin na rośliny jest silna stymulacja wzrostu międzywęźli prowadząca do wydłużania się pędów. Przy pomicy giberelin można przywrócić normalny wzrost i pokrój karłowatym mutantom. Hormony te mogą również wywoływać kwitnienie u roślin wymagających do zakwitnięcia długich dni. Gibereliny działają również antagonistycznie do auksyn, ponieważ pobudzają do rozwoju pąki. Wywołują one również partenokarję, czyli rozwój owoców baz zapłodnienia (np. grusza). Odgrywają one również dużą rolę w procesie przerywania stanu głębokiego spoczynku utworów przetrwalnikowych (nasion), mogą one w tym procesie zastąpić niektóre czynniki jak chłód i światło.
31.ZNACZENIE FOTOINDUKCJI I TERMOINDUKCJI W PROCESIE WZROSTU I ROZWOJU ROŚLIN.
Światło jest czynnikiem silnie wpływającym na morfogenezę rośliny. Pęd siewki rozwijający się w ciemności jest blady i wiotki. Pozbawiony jest chlorofilu, hipokotyl i międzywęźla są wydłużone, blaszki liściowe zredukowane, wierzchołek zgięty w dół. Zjawisko to nosi nazwę etiolacji (wypłowienia). Na świetle pęd zazielenia się, łodyga skraca, blaszki liściowe są dobrze rozwinięte, wierzchołek wyprostowany. Cechy odróżniające rozwijającą się w ciemności od rośliny rozwijającej się w świetle to fotomorfozy. Ogromna rola światła w rozwoju roślin jest związana z fotosyntezą, lecz znaczenie światła nie ogranicza się do fotosyntezy i produkcji asymilatów, ponieważ zmiany w rozwoju siewki pod wpływem światła nie zależą od fotosyntezy. Dowodzi tego bardzo niskie stężenie światła lub bardzo krótki czas naświetlenia jaki wystarcza do wywołania fotomorfozy. W przypadku wypłoweiałych siewek wystarczy pojedynczy błysk słabego światła, aby wywołać po 10 minutach spowolnienie wzrostu łodygi, zapoczątkowanie wyprostowania się wierzchołka pędu i syntezy chlorofilu. Światło działa tu nie jako źródło energii, ale jako sygnał do wywołania zmian rozwojowych. Nosi to nazwę FOTOINDUKCJI. Inne fotomorfozy to: zmiana kształtu komórek szparkowych ( wraz z otwieraniem się szparek), kiełkowanie niektórych nasion. Indukcja biegunowości w komórkach, różnicowanie się przedrośla u paproci, synteza karotenoidów i wzrost sporangioforów u grzybów. Fotomorfozy zachodzą pod wpływem światła czerwonego i niebieskiego.
U wielu roślin konieczne jest działanie niskich temperatur, w celu zainicjowania kwitnienia, lub przynajmniej niska temperatura kwitnienie przyspiesza . Działanie chłodu ma tu charakter bodźcowy i nazywa się TERMOINDUKCJĄ. Indukowany niską temp. Umożliwiający kwitnienie nosi nazwę wernalizacji lub jaryzacji. Termoindukcja zachodzi pod wpływem temp. Od nieco poniżej zera do około 10oC. Wymagany jest przy tym dłuższy czas działania niskiej temp. co najmniej 3-8 tygodni. Gdy w tym czasie proces wernalizacji zostanie przerwany przez podwyższenie temp., efekt termoindukcji nie pojawi się (dewernalizacja). Wrażliwość rośłin na okresowe działanie chłodu jest różna: od niewrażliwości ( u roślin jednorocznych ), poprzez przyspieszenie kwitnienia, do pełnej zależności kwitnienia od termoindukcji. Rośliny ozime przechodz ą wernalizację w stadium spęczniałego nasienia. Rośliny 2-letnie stają się wrażliwe na wernalizację po osiągnięciu pełnego rozwoju wegetatywnego.
Miejscem percepcji bodżca wernalizacyjnego jest merystem wierzchołkowy pędu. W laboratorium można chłodźić same wierzchołki pędów i doprowadzać rośliny do kwitnienia. U roślin 2-letnich wymagających wernalizacji można ją zastąpić podaną z zewn. gibereliną, która wymagana jest głównie do „wystrzelenia” wysokiego pędu kwiatonośnego ( rośliny rozetkowe).
32.ZNACZENIE N I Mg W METABOLIŹMIE
Azotu w tkankach roślinnych jest dużo, do 6% suchej masy. Zajmuje on 4 miejsce pod względem zawartości w biomasie po C, O, H.
Azot jest pierwiastkiem niezbędnym do budowy aminokwasów, a także białek, nukleotydów i kwasów nukleinowych (biorących udział w metabolizmie). Bierze udział prawie we wszystkich reakcjach biochemicznych. Dlatego niedobór N stanowi częsty czynnik ograniczający w tworzeniu nowych tkanek. Metabolizm azotowy jest ściśle powiązany z przemianami węgla. Głównymi formami azotu dostępnymi dla roślin są jony NO3- i NH4+ . Pobrane przez roślinę azotany muszą ulec redukcji do formy NH2 , gdyż w takiej postaci są wbudowane w aminokwasy i białka. Proces redukcji wymaga energii płynącej z oddychania. Obecność azotanów lub soli amonowych wpływa na pobieranie innych pierwiastków. Jony amonowe stanowią większość kationów pobieranych przez roślinę. Pobieranie innych kationów zostaje zmniejszone, natomiast pobieranie anionów dla zachowania równowagi zwiększone. W przypadku azotanów jest odwrotnie, gdyż stanowią one większość pobieranych anionów.
Pobieranie MAGNEZU następuje w postaci jonów Mg2+ . Rola Mg związana jest ze zdolnością tworzenia wiązań chelatowych (związek chelatowy to połączenie metalu z cząsteczką organiczną). Zadzniem chelatów w glebie jest utrzymanie metali w formie przyswajalnej, metale w postaci chelatów są całkowicie dostępne dla roślin. Mg bierze udział w przekształcaniu, przenoszeniu i magazynowaniu energii. Umożliwia właściwy przebieg reakcji glikolizy, syntezy materiałów budulcowych i zapasowych, fosforylacji fotosyntetycznej i oksydacyjnej. Występuje w:
-chloroplastach i rybosomach (chlorofil jest 4-kleszczowym chelatem Mg)
-wiązaniach chelatowych
Brak Mg hamuje fotosyntezę, oddychanie, a także gromadzenie węglowodanów, białek i tłuszczów w roślinie.
33.OGóLNA CHARAKTERYSTYKA I ZNACZENIE PROCESÓW KATABOLICZNYCH I ANABOLICZNYCH
Funkcjonowanie żywej komórki( przemiana materii ) , jej wzrost , podziały i różnicowanie , a także rozwój całego organizmu są wynikiem precyzyjnej organizacij strukturalno-funkcjonalnej i jej regulacji. Przemiany metaboliczne stanowią integralną część tej organizacji, a regulacja procesów metabolicznych - podstawowy element regulacji procesów życiowych. Zmiany w wartości składników organicznych są wynikiem 2-óch rodzajów przemian, przebiegających w innych kierunkach. Przemiany, w których z prostych związków powstają nowe, bardziej skomplikowane substancje organiczne, nazywamy anabolizmem (procesy syntezy). Natomiast procesy degradacji (stopniowego rozkładu) bardziej złożonych substancji do związków prostych nazywamy przemianami katabolicznymi. W wyniku tych procesów związki organiczne przekształcają się ostatecznie w substancje nieorganiczne. W przypadku większości składników ,których zawartość nie zmienia się widocznie, odbywa się ciągła ich synteza i degradacja. Obserwowane stałe stężenie jest więc wynikiem stanu równowagi dynamicznej między przemianami anabolicznymi a katabolicznymi tego składnika. Intensywność tych przemian określamy jako szybkość odnowy składnika.
Procesem anabolicznym jest fotosynteza. Jest ona najważniejszym procesem zachodzącym w roślinach zielonych. W procesie tym energia słoneczna zostaje zamieniona na energię chemiczną związków organicznych. Prawie wszystkie związki org. znajdujące się na ziemi powstały w procesie fotosyntezy. Sumaryczne równanie fotosyntezy:
CO2+2H2O=====C6H12O6+O2+H2O
Fotosyntezie towarzyszy charakterystyczna wymiana gazowa i pobieranie CO2 i wydzielanie tlenu. Rezultatem produkcji związków organicznych przez fotosyntezę jest przyrost suchej masy rośliny.
Mianem fotosyntezy obejmuje się całość procesu prowadzącego do redukcji CO2 , choć sama redukcja CO2 nie wymaga swiatła.
Wyróżniono 2 fazy fotosyntezy:
faza reakcji fotochemicznych, wymagająca światła , wynikiem której jest redukcja NADP oraz synteza ATP.
faza reakcji enzymatycznych, nie wymagająca światła, lecz zużywająca produkty reakcji fotochemicznych ( NADPH , ATP ). W tej fazie następuje związanie i redukcja CO2.
W procesie fotosyntezy światło musi zostać zaabsorbowane. Służą temu barwniki fotosyntetyczne:
chlorofile , karetonoidy , fikohiliny
Oddychanie jest procesem katabolicznym. Jest to proces dysymilacji w którym następuje rozkład substancji organicznych z powrotem do H2O i CO2. Oddychaniu towarzyszy charakterystyczna wymiana gazowa-
wydzielaniu CO2 towarzyszy pobieranie O2. Następuje przy tym uwolnienie energii. Substraty oddechowe to przede wszystkim cukrowce, ale tez tłuszcze. U roślin spotykamy pewne modyfikacje tego procesu, które go różnią od oddychania zwierząt. Równanie sumaryczne:
C6H12O6+6O2+2H2O===6CO2+12H2O
Uwalnia się przy tym energia , której część pojawia się w ATP. Główne etapy oddychania to glikoliza , cykl kwasów trikarboksylowych i oddechowy łańcuch transportu elektronów.
34. MECHANIZM TRANSPORTU WE FLOEMIE
Podobnie jak w przypadku transportu ksylemowego, podstawą oceny, jakie związki i jony oraz w jakich ilosciach są transportowane przez floem, jest analiza chemiczna soku floemowego( na podstawie soku wypijanego bezpośrednio z rurek sitowych przez mszyce) Skład chemiczny soku floemowego jest cechą gatunkową ,nie zawiera cukrów redukujących). Przez tą tkanke przewodzącą w czasie starzenia się liści następuje export dużych ilości produktów rozkładu białek w chloroplastach i mitochondriach, zaopatrzenie organów generatywnych w organiczne związki azotowe przemieszczane z organów wegetatywnych. Przez rurki nitowe przemieszczają się wszystkie hormony. We floemie występują również kwasy organiczne, witaminy oraz ATP i ADP
Na przekroju poprzecznym łodygi rurki sitowe stanowią ok. 1/5 pow łyka o sprawnosci transportu floemowego decyduje liniowa szybkość przepływu , jak tez stężenie przemieszczanych związków w soku floemowym. Intensywność transportu;
L=C*V[g*cm-2*h-1]
L-masa transportu substancji w jednostce czasu;
stężenie(g*cm-3)
V-szybkość liniowa(cm*h-1)
Intensywność transportu floemowego jest różna dla różnych gat i mniejsza od liniowej intensywności transportu ksylemowego.
Przeciętnie wynosi ona około 0,6-5 g*cm-2*h-1
Czynniki wpływające na transport floemowy:
-temperatura i zmienność dobowa :
a)temp w zakresie ok. 50st C słabo wpływa na ten proces
b)temp ma wpływ na intensywność rozładunku, słabo wpływa na przepływ przez rurki sitowe.
„Siłownia” floemu (centrum syntezy ATP) są komórki towarzyszące.
Transport asymilatów odbywa się przez całą dobę z różną intensywnością , co świadczy o braku zależności tego procesu od fotosyntezy.
Mechanizm transportu floemowego na duże odległości:
załadunek floemu może się odbywać w dwojaki sposób;
przez symplastyczny kontakt-plazmodesmy między komórkami miękiszowymi a komórk towarzyszącymi i rurkami sitowymi
na drodze apoplastycznej
rozładunek floemu- odbywa się w komórkach akceptorów do których transportowane są substancje przemieszczane przez floem (przebiega to zgodnie z malejącym gradientem potencjału chemicznego)
Sprawność rozładunku floemu i metabolizm substancji transportowych determinuje wielkość importu substancji pokarmowych do akceptorów
Rozładunek może odbywać się przez symplast i apoplast:
akceptory intensywnie rosnące (młode liście) zaopatrywane są przez symplast
akceptory akumulujące (organy spichrzowe) importują zw węglowe przez apoplast
W organach generatywnych, w rozwijajacych się nasionach, u których brak jest kontaktu symplastycznego między kom zarodka a kom rośliny macierzystej , rozładunek jest apoplastyczny.
Siłą motoryczną ruchu w rurkach sitowych jest gradient ciśnienia hydrostatycznego, czyli turgorowego, miedzy punktem załadunku i rozładunku floemu. W teorii przepływu masowego związki i jony przemieszczają się w rurkach sitowych pod ciśnieniem wraz z wodą, w każdej rurce tylko w jednym kierunku. Gradient potencjału wody między rurami sitowymi i ksylemem powoduje stałą cyrkulację wody.
35. ZNACZENIE P , WYSTĘPOWANIE.
P
Występowanie
- związki organiczne ( kwasy nukleinowe, fosfolipidy)
-związki nieorganiczne ( pochodne kwasu fosforowego, związki zawierające Ca, Fe lub Al.)
Pobieranie;
- w postaci jonów kwasu ortofosforowego (postać utleniona)
Przyswajalność zależy od pH ( pH<5 - P tworzy z Fe Ca i Al. nierozpuszczalne związki.) Najlepsza przyswajalność 6<pH<7. Fosfor ze związków organicznych jest dostępny dopiero po ich mineralizacji.
Występowanie w roślinach
- nukleotydy (magazynujące i przenoszące energię -ATP, transportujące wodór w procesach oksydoredukcyjnych- NAD , NADP , FAD , przenoszące grupy czynne)
- kwasy nukleinowe DNA, RNA.
- estry fosforanowe cukrów
- fosfolipidy
-zapasowe związki fosforu np. fityna.
36 .MIEJSCE SYNTEZY , DROGI I MECHANIZMY TRANSPORTU AUKSYNY.
Biosynteza auksyny ( kwasu inohilo-3- octowego IAA) odbywa się w rosnących tkankach ( wierzchołki wzrostu, młode liście, pąki , kwiaty, owoce) z aminokwasu aromatycznego ( tryptofanu) w procesach deaninacji , dekarboksylacji i utleniania. Auksyna jest aktywnie transportowana przez kambium bazypetalnie czyli od końca apikalnego w kierunku bazypetalnego. Kierunek transportu jest związany z biegunowym zróżnicowaniem komórek. O polarnym transporcie auksyny świadczy doświadczenie w którym w pędzie wierzbowym wycięto pierścień kory wraz z łykiem przecinając tym samym drogę aktywnego transportu auksyny. Po nałożeniu na koniec apikalny auksyny, po pewnym czasie okazało się ,że auksyna wywarła hamujący wpływ narozwój pączków śpiących tylko do strefy przecięcia drogi transportu. W dolnej części pędu pąki rozwinęły się w pędy- auksyna nie zadziałała. O transporcie auksyny tylko w kierunku bazypetalnym świadczy doświadczenie w którym auksynę nałożono na koniec bazalny. Nie została ona przetransportowana aktywnie, tym samym nie zahamowała rozwoju pąków śpiących które rozwinęły się w pędy. Szybkość transportu polarnego auksyny wynosi około 10cm /h. Transport tego hormonu nie polega na przemieszczaniu się jej od korony wzdłuż całego pnia. Auksyna jest bodźcem indukującym szereg łańcuchowych reakcji w komórkach wrzecionowatych kambium. Pobudza ona komórki do intensywnych podziałów.
CZYNNIKI WARUNKUJĄCE INTENSYWNOŚĆ FOTOSYNTEZY U DRZEW LEŚNYCH.
Na efektywność fotosyntetyczną wpływają różne czynniki środowiska. Najważniejsze z nich to: woda CO2,światło, temperatura.
WODA - jej wpływ nie jest bezpośredni, ponieważ zawartość H2O w chloroplastach dla potrzeb fotosyntezy jest wystarczająca.
Jednak wpływa ona silnie na asymilację CO2, gdyż w przypadku deficytu wody zamykają się aparaty szparkowe i praktycznie zostaje odcięty dopływ CO2 z powietrza do liści, prowadząc do spadku stężenia CO2 w przestworach międzykomórkowych. Ponadto jej niedobór hamuje wzrost roślin i w wyniku zmniejszenia blaszek liściowych mniej światła i CO2 jest odbierane.
ŚWIATŁO-
świetlny
punkt
kompensacyjny
Krzywa świetlna obrazuje zmiany natężenia fotosyntezy , mierzone ilością pobranego CO2 pod wpływem różnych natężeń światła.
Przy bardzo niskich natężeniach światła, mimo że fotosynteza się odbywa nie obserwujemy pobierania CO2 , a jedynie zmniejszona jego wydalanie.
Równocześnie z fotosyntezą odbywa się oddychanie, które zużywa tlen i wytwarza CO2. Początkowo fotosynteza zużywa CO2 powstały w procesie oddychania, nie pobierając go z atmosfery. Przy zwiększonym natężeniu światła wymiana gazowa ustaje - brak pobierania lub wydalania CO2 orazO2 . takie stężenie światła w którym dochodzi do zrównania fotosyntezy i oddychania oraz ustania wymiany gazowej to świetlny punkt kompensacyjny. Jego wartość zależy od gatunku i warunków rozwoju drzewa.
Przy ilościach światła powyżej punktu kompensacyjnego następuje liniowy wzrost pobierania CO2, co jest miarę natężenia fotosyntezy netto.
Wzrost ustaje gdy ilość światła osiąga określoną wartość. Powyżej określonego stężenia nie następuje wzrost fotosyntezy. Jest to świetlny punkt wysycenia. Jego wartość zależy od gatunku i warunków rozwoju.
Bardzo wysoki stopień natężenia światła przy długim czasie naświetlania może hamować fotosyntezę poprzez uszkodzenie aparatu asymilacyjnego.
CO2-
CO2
μmol*m-2*s-1
(fotosynteza)
30
25
20 stężenie
15 kompensacyjne
10
5
0 100 200 300 CO2 m/l
-5
Wpływ stężenia CO2 na fotosyntezę, której miarą jest stężenie CO2 w jednostce czasu na jednostkę powierzchni liścia. Poczynając od bardzo niskich wartości stopniowe zwiększenie CO2 powoduje wpierw dojście fotosyntezy do stanu równowagi między fotosyntezą a oddychaniem, stężenie w którym ten stan się pojawia nazywamy stężeniem kompensacyjnym , a następnie wzrost natężenia fotosyntezy.
Stężenie CO2 o wartości 0,1% jest optymalna dla rośliny, a 1% już trujące. Korzystne jest większe stężenie CO2 przy większej temperaturze i natlenieniu.
TEMPERATURA-
CO2
μmol*m-2*s-1
(fotosynteza)
50
40 silne natężenie
30 światła
20
10 słabe natężenie światła
0
0 10 20 30 40
°C
Wpływ temperatury na asymilację CO2 przy silnym i słabym natężeniu światła.
Przy niskim natężeniu światła temperatura w zakresie do 30°C nie ma wpływu na fotosyntezę -czynnikiem ograniczającym jest wtedy światło. W wyższych temperaturach natężenie fotosyntezy zmienia się (nasila się fotorespiracja + ogólnie hamujący wpływ temperatury na efektywność wielu enzymów)
Temperatura może być czynnikiem ograniczającym, zwłaszcza w warunkach wysycenia aparatu fotosyntetycznego światłem (powyżej świetlnego punktu wysycenia)przy dobrym zaopatrzeniu w CO2 .
Przy wysokim stężeniu światła krzywa zależności fotosyntezy od temperatury ma wyraźne optimum około 30°C jest to wartość charakterystyczna dla reakcji enzymatycznych.
1999
Strona 1 z 29
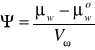
Szybkość
reakcji
Szybkość reakcji
Natężenie światła
Świetlny punkt
wysycenia
Pobieranie
CO2
Wydalanie
CO2
Wyszukiwarka